
Coptobrycon bilineatus ( Ellis, 1911 )
|
publication ID |
https://doi.org/ 10.1590/S1679-62252010000400004 |
|
persistent identifier |
https://treatment.plazi.org/id/2B6EF01C-FFDF-7553-FC07-FE12FBFE6DDD |
|
treatment provided by |
Carolina |
|
scientific name |
Coptobrycon bilineatus ( Ellis, 1911 ) |
| status |
|
Coptobrycon bilineatus ( Ellis, 1911) View in CoL
Figs. 1-7 View Fig View Fig View Fig View Fig View Fig
Hasemania bilineata Ellis, 1911: 150-151 View in CoL (type locality: “Alto da Serra, São Paulo, in a creek” - São Paulo, upper rio Tietê basin); Eigenmann, 1921: 223-224 (copy of original description); Böhlke, 1958: 45 (compared to Pristicharax hanseni View in CoL ); Géry, 1966: 226-228 (transfer to Coptobrycon View in CoL ); Géry, 1977: 378 and 402 (key, diagnosis and illustration).
Coptobrycon bilineatus View in CoL . - Géry, 1966: 224-235 (rio Tietê basin, Brazil); Géry, 1972: 9 (citation); Lima & Gerhard, 2001: 111- 112 (citation); Géry, 1977: 378 and 402 (key, diagnosis and illustration); Serra et al., 2007: 132, 134, 135, table 1, fig. 3b (rio Itatinga, São Paulo State coastal drainage, RPPN Parque das Neblinas; biogeography; conservation; ecology); Serra et al., 2008: 60-61 (conservation status, ecology, distribution).
Material examined. Brazil. São Paulo. Bertioga. Rio Itatinga, RPPN Parque das Neblinas. DZSJRP 6601 , 4 , 17.0- 26.7 mm SL, near the Visiting Center, at the footbridge, 23º44’55”S 46º09’44”W. DZSJRP 6610 , 150 , 18.4-36.7 mm SL, and DZSJRP 6889 , 2 c&s, 27.9 and 36.3 mm SL, under concrete bridge, 2 km from the Visiting Center, 23º44’47.2”S 46º09’29.7”W. DZSJRP 6614 , 13 , 26.7-33.7 mm SL, ca. 9 km from the Visiting Center, 23º47’42”S 46º12’14”W. DZSJRP 6621 , 94 , 25.7-36.7 mm SL, and DZSJRP 6890 , 2 c&s, 28.1 and 36.3 mm SL, mouth of tributary creek on the right bank of the rio Itatinga, fifth bridge on dirt road from the Visiting Center, 23º47’17.7”S 46º11’26.0”W. DZSJRP 6624 , 1 , 27.2 mm SL, mouth of tributary creek on the right bank of the rio Itatinga , fourth bridge on dirt road from the Visiting Center, 23º46’52.6”S 46º10’56.2”W. DZSJRP 6626 , 3 , 23.2-28.4 mm SL, mouth of tributary creek on the right bank of the rio Itatinga , third bridge on dirt road from the Visiting Center, 23º46’21.2”S 46º10’11.5”W. DZSJRP 6631 , 10 , 25.7-32.1 mm SL, mouth of tributary creek on the right bank of the rio Itatinga , second bridge on dirt road from the Visiting Center, 23º45’47.6”S 46º09’41.0”W. DZSJRP 6888 , 1 c&s, 19.3 mm SL, rio Itatinga , near the Visiting Center, at the footbridge, 23º44’55”S 46º09’44”W. Santo André. MZUSP 4516 View Materials , 4 View Materials , 1 View Materials c&s, 26.6-30.0 mm SL, Campo Grande, near Paranapiacaba GoogleMaps .
Diagnosis. The same as for the genus.
Description. Morphometric data in Table 1. Small size, largest specimen examined 37 mm SL. Body moderately high, greatest body depth slightly ahead vertical through dorsal-fin origin. Dorsal profile of head straight or slightly convex from tip of snout to posterior portion of supraoccipital spine. Dorsal profile of body convex, from supraoccipital spine to dorsalfin origin; straight, or slightly convex, and descending along dorsal-fin base; straight and gradually descending from dorsal fin to caudal peduncle end. Head rounded in transverse
F. Langeani & J. P. Serra 729
section; predorsal portion slightly keeled mainly on posterior half; postdorsal portion slightly flattened anteriorly and slightly keeled posteriorly.Ventral profile of body convex from snout to pelvic-fin origin; straight and horizontal from pelvic fin to anal-fin origin (considerably more convex in specimens with replete digestive tube); straight, or slightly concave, and ascending along anal-fin base; straight and slightly ascending, nearly horizontal, from anal fin to caudal-fin base. Ventral portion of head and body up to anal-fin origin rounded in transverse section; base of anal fin keeled; caudal-peduncle ventral portion rounded, slightly keeled near anal-fin base.
Head triangular from lateral view; snout rounded. Mouth terminal. Premaxilla short; maxilla short, extending to behind vertical passing through anterior margin of orbit. Anterior and posterior nasal pores completely separated ( Fig. 2 View Fig ); anterior nasal pore slightly elevated, on apex of short cone, and posterior pore not elevated, at same level of snout surface; olfactory rosette with up to six lamellae, posteriorly directed and turned to posterior nasal pore ( Fig. 3 View Fig ). Adipose eyelid underdeveloped, mostly inconspicuous, except in anteroventral portion of eye.
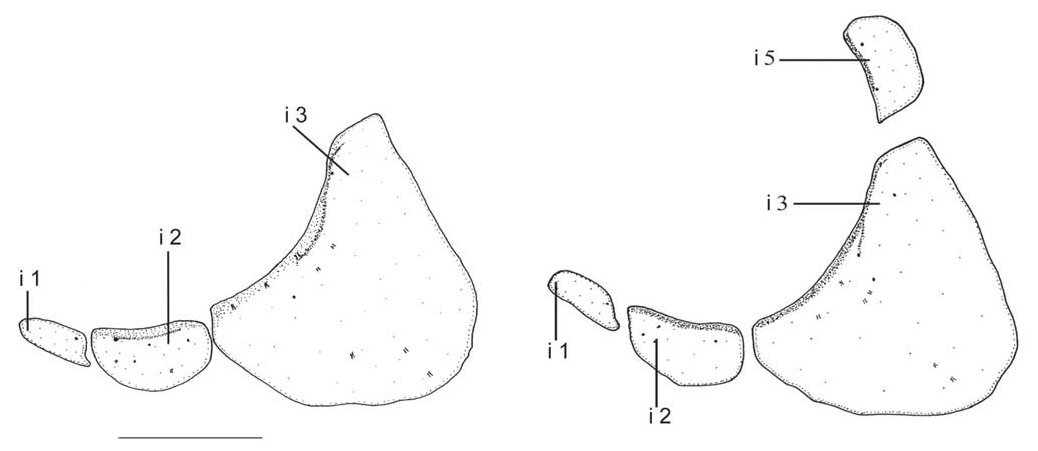
Frontoparietal fontanel from posterior portion of mesethmoid to base of supraoccipital spine, visible and conspicuous in c&s specimens, but not apparent and covered by thick skin in alcohol-preserved specimens. Rhinosphenoid absent, only cartilaginous tissue present between lateral ethmoid and vomer. Orbitosphenoid with posterior invagination limiting ventral projection, posterior and elongated, turned to pterosphenoid. Fourth and sixth infraorbitals absent; fifth infraorbital also absent in some specimens ( Fig. 4 View Fig ); when present, separated from third infraorbital by space equivalent to half its depth ( Fig. 4 View Fig ). Nasal and supraorbital absent; antorbital vertically elongated with anteroventral process crossing anterior process of maxilla and turning to premaxilla dorsoposterior portion.
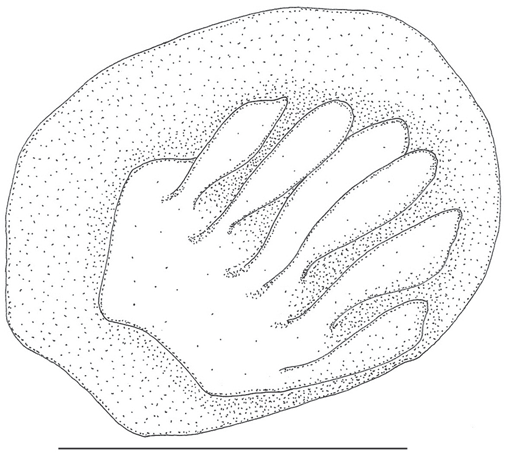
Canals and pores of cephalic lateral line present and well developed along latero-anterior portion of frontal (extending only up to vertical passing through epiphyseal bar), behind anterior nasal pore, on dentary and preopercle; infraorbital bones with canals underdeveloped or absent and inconspicuous pores. Free neuromasts (sensory papillae) present along the supraorbital, infraorbital, maxilla, dentary, gular, and supraoccipital cephalic laterosensory series.
Four branchiostegal rays. Gill rakers elongated on first branchial arch; two on hypobranchial, eight to ten on ceratobranchial and five to six on epibranchial.
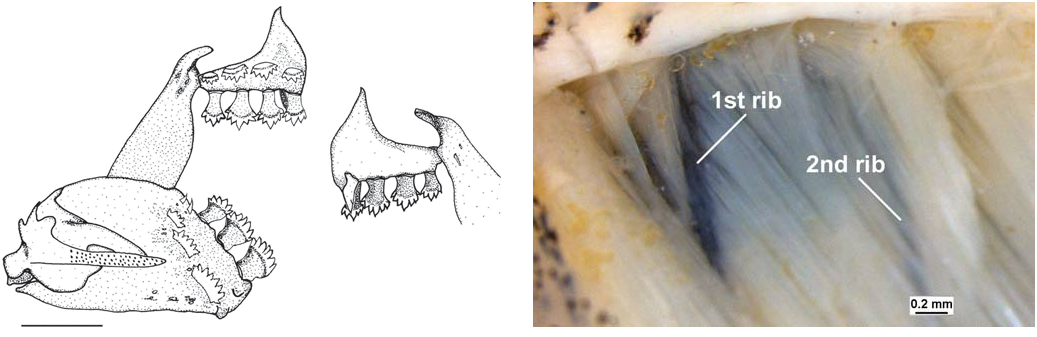
Outer row of premaxilla with one tri- to tetracuspid tooth (rarely with more than four cusps), between first and second teeth of inner row and slightly turned anteriorly; inner row with four (33) or five (2) pedunculated, penta- or octocuspid 730 Redescription and relationships of Coptobrycon bilineatus teeth, with wide compressed crown, slightly curved posteriorly, with median cusp subequal, slightly more developed than others ( Fig. 5 View Fig ). Maxilla without teeth. Dentary short, with single row of two (2), three (31) or four (2) anterior, large, hepta- to decacuspid, pedunculated teeth, with large, compressed, straight and turned posteriorly crown, with rounded subequal cusps, followed by one to four much smaller conical teeth ( Fig. 5 View Fig ). Palatine approximately rectangular, without openings, perforations or thinning in its mid-anterior portion.
Hypaxialis muscle portion largely reduced, especially obliquus superioris muscle, leaving thin muscular layer, composed principally of obliquus inferioris, in front of first pleural rib, exposed anterior and proximally ( Fig. 6); muscle reduction also between first and second pleural ribs. No direct contact of swim bladder with skin.
Scales cycloid and relatively large, similar in size to diameter of pupil. Lateral line incomplete, with only two (2), three (20), or four (7) perforated scales, and 26(1), 27(3), 28(6), 29(8), 30(9), 31(1), or 32(2) non-perforated scales. Five (5) or six (26) longitudinal rows of scales above lateral line and four (28) or five (3) below. Two or three scales slightly covering base of first anal-fin rays, visible with high magnification. Twelve (21) or 14(11) scales around caudal peduncle. Predorsal line with regular row of 10(2), 11(4), 12(7), or 13(6) scales. Thirty-two (2), 33 (2) or 34 (1) vertebrae.
Dorsal fin with ii,7(3) or ii,8(30) rays; with posterior rounded edge, in posterior half of body, little behind vertical passing pelvic-fin origin; first unbranched ray almost one-third or half longer than second unbranched one, slightly shorter than first branched ray.Adipose fin absent. Pectoral fin small, with i,8(1), i,9(5), i,10(25), or i,11(2) rays; with posterior rounded edge, not reaching pelvic-fin base, its posterior extremity beyond vertical passing through beginning of last third of distance between its base and pelvic-fin base.
Coracoid little developed ventrally. Opening between coracoid and cleithrum nearly entirely visible laterally; scapula welldeveloped,globose, withsmallscapularforamen, morecentral and bordered anteriorly by wide bony portion ( Fig. 7 View Fig ). Pelvic fin with i,4(1), i,5(28) or i,6(3) rays; with posterior rounded edge, not reaching anal-fin origin; its posterior extremity beyond vertical passing through last third of distance between its origin and anal-fin origin; anal fin with iii,10(1), iii,11(19), or iii,12(13) rays; with straight or slightly concave edge; second unbranched ray and two first branched rays, longer and subequal, subsequent rays gradually shorter.Caudal fin with rounded lobes and i,17,i(33) principal rays.Mature males and females (dissected from DZSJRP 6610 and 6621) without hooks on pelvic and anal-fin rays.
F. Langeani & J. P. Serra 731
Color in life. Body predominantly yellowish ventrally. In water, specimens appear golden, probably as a consequence of great quantities of guanine along portions of visceral cavity, infraorbital and opercular regions; lateromedial and lateroventral black conspicuous stripes. Fins yellowish orange, especially dorsal, anal, and caudal fins.
Color in alcohol. Black chromatophores distributed all over body. Head with large concentration of small chromatophores along lateral portion; larger chromatophores on dorsal portion of operculum and below eye, on third infraorbital; regions of larger chromatophores bordered by clear areas, without pigment. Concentrated chromatophores near edges of scales, forming reticulate pattern. Humeral spot absent; mid-lateral stripe from vertical passing dorsal-fin origin, or posterior portion of head (more melanic specimens), up to caudal-fin base; shorter stripe, slightly above anal-fin base, from analfin origin, or little behind, through ventral portion of caudal peduncle up to caudal-fin base; caudal-fin base dark. Dorsal portion of body dark from head to caudal fin; head chromatophores small and numerous; from head to dorsal-fin origin as in rest of body; posterior to dorsal fin much more numerous, larger, conspicuous, and visible laterally. Ventral portions of head and body lighter, with small and numerous chromatophores along dentary, anterior portion of isthmus, and ventral portion of preopercle and interopercle; chromatophores slightly larger and more scattered from posterior portion of head to pelvic-fin origin, and numerous, dense and conspicuous from anal pore to caudal fin. Fins slightly dark, mainly along anterior and posterior edges of rays.
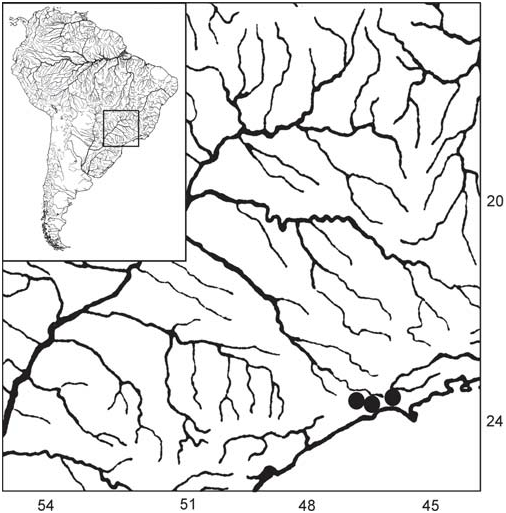
Distribution. Upper rio Tietê (upper rio Paraná drainage) and rio Itatinga, a tributary of the rio Itapanhaú, a small independent coastal river system, São Paulo State, Brazil ( Fig. 8 View Fig ).
Ecology and conservation. The RPPN Parque das Neblinas is a privately owned conservation unit within the Atlantic Forest, and most of the rio Itatinga basin is within the boundaries of this protected area. The rio Itatinga ( Fig. 9 View Fig ) is a typical Atlantic Forest stream, possessing swift-flowing, transparent, cold waters, and alternating relatively deep pools and shallow riffles. Usually, the bottom of the more lotic portions is composed of sand, stones and boulders, whereas in the more lentic stretches there is an accumulation of leaves and branches from the riparian vegetation. The riparian vegetation is an old, abandoned eucalyptus plantation, with dense undergrowth of native shrubs and trees.
Coptobrycon bilineatus was common and abundant in most of the sampled stretches of the rio Itatinga. Specimens were easily observed forming large schools, often associated with schools of Astyanax paranae . The species was generally recorded in areas with a relatively slow water flow, among macrophytes, branches and submerged logs. The examination of some stomach contents revealed mainly fragmented terrestrial vegetation, although some digested items of animal origin were recorded in smaller quantities. Immature, maturing, mature, and empty gonads were recorded, which seems to indicate that the species reproduces in more than one season of the year. Other species collected at the rio Itatinga are Astyanax paranae , Glandulocauda melanogenys [= G. melanopleura Ellis, 1911 sensu Menezes & Weitzman, 2009 ], Taunaya bifasciata , Trichomycterus sp. [= T. iheringi (Eigenmann, 1917) ], Pseudotocinclus tietensis , and Phalloceros caudimaculatus [= P. reisi Lucinda, 2008 ] ( Serra et al., 2007).
Until recently, Coptobrycon bilineatus was considered to be endemic to the upper course of the rio Tietê ( Langeani, 1989), where it was collected for the last time in 1981 (ZUEC 4287). Extensive collecting in the upper rio Tietê basin, conducted from 1985 to date was not successful in finding any additional specimens of the species. Coptobrycon bilineatus is officially considered an endangered species in Brazil ( Serra et al., 2008). The discovery of Coptobrycon bilineatus in the rio Itatinga is, from a conservation point of view, quite reassuring. However, its overall conservation status is still cause for concern and special attention to the species is required.
732 Redescription and relationships of Coptobrycon bilineatus




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Coptobrycon bilineatus ( Ellis, 1911 )
| Langeani, Francisco & Serra, Jane Piton 2010 |
Coptobrycon bilineatus
| Serra, J & Langeani, F 2008: 60 |
| Serra, J 2007: 132 |
| Gery, J 1977: 378 |
| Gery, J 1972: 9 |
| Gery, J 1966: 224 |
Hasemania bilineata
| Gery, J 1977: 378 |
| Gery, J 1966: 226 |
| Bohlke, J 1958: 45 |
| Eigenmann, C 1921: 223 |
| Ellis, M 1911: 151 |