
Farlowella yarigui, Ballen, Gustavo A. & Mojica, José Iván, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3765.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:3E5C83F5-508F-41F2-8B3F-1A0C3FD035FE |
|
DOI |
https://doi.org/10.5281/zenodo.5673229 |
|
persistent identifier |
https://treatment.plazi.org/id/275FA33D-FFFC-FFB3-FF7F-2332962C2D20 |
|
treatment provided by |
Plazi |
|
scientific name |
Farlowella yarigui |
| status |
sp. nov. |
Farlowella yarigui View in CoL , new species
( Figure 1 View FIGURE 1 , Table 1 View TABLE 1 )
Holotype. ICNMHN 17819, adult male, 112.3 mm SL, COLOMBIA, Departamento de Santander, El Carmen de Chucurí, Vereda El Topón, río Topón, main channel of the river, 6°42’00” N, 73°32’31” W, 422 masl, 19 March 2011, J. I. Mojica, G. A. Ballen, and biology students, Universidad Nacional de Colombia.
Paratypes. COLOMBIA, Departamento de Santander, El Carmen de Chucurí, Vereda El Topón, río Topón, ICNMHN 18889 (same data as the holotype), 2, 36.2–43.4 mm SL; río Topón, main channel, 6°45'45"N, 73°34'56"W, 276 masl, 18 March 2011, J. I. Mojica, G. A. Ballen, and biology students, Universidad Nacional de Colombia, ICNMHN 17789, 2, 75.6–76.9 mm SL.
Diagnosis. Farlowella yarigui differs from species of the F. curtirostra species group by having restricted odontodes on sides of the snout but not extending onto the cheek region (vs. hypertrophied odontodes in breeding males extensive on sides of the snout and cheek in species of the F. curtirostra species group). In addition, F. yarigui differs from species of the F. amazona species group by presenting notably concave lateral margins of the snout (vs. lateral margins of the snout straight to nearly straight in the F. amazona species group); and eyes not elevated on the head (vs. eyes elevated on the head in the F. amazona species group). Farlowella yarigui differs from species of the Farlowella nattereri species group as well as from Farlowella gracilis by presenting four rows of lateral plates (vs. five rows in F. gracilis and the F. nattereri species group). In addition, Farlowella yarigui differs from species of the F. knerii and F. mariaelenae species groups by presenting two abdominal plate rows (vs. three abdominal rows in the F. knerii and F. mariaelenae species groups).
Farlowella yarigui is herein assigned to the Farlowella acus species group by the presence of long breeding odontodes on the snout in mature males; four rows of lateral body plates from the anteriormost portion of body to posterior margin of dorsal- and anal-fin base; two complete rows of ventral plates and no single plate between pelvic-fin insertions. Farlowella yarigui differs from all the species currently assigned to the F. ac u s species group by having the median anterior lateral row of plates diamond-shaped (vs. hexagonal in shape in F. a c u s, F. colombiensis , F. martini , F. venezuelensis and F. vittata . In addition, Farlowella yarigui differs from F. martini by having dark interradial membranes in the caudal fin (vs. interradial membranes clear in F. martini )
Description. Largest specimen 16.8 mm SL (adult male, holotype). Overall shape elongate and cylindrical. Head slightly depressed, body cylindrical, tail very depressed. Dorsal profile from tip of snout to level of nares concave, almost entirely straight from level of dorsal-fin insertion to caudal peduncle. Ventral profile obliquely straight from tip of snout to pectoral girdle and then straight to caudal peduncle. Body completely covered with plates except for tip of snout and gular region.
Head in dorsal view with concave lateral margins from snout to halfway between tip of snout and nares, and gently concave from such landmark to opercular region. Snout short with papillary naked tip, papillae more evident in adults; hypertrophied odontodes present on sides of snout in mature males but restricted to snout and not reaching sides of head. Preorbital ridge present; nares with anterior opening smaller than posterior opening, dermal flap separating both openings present. Eyes lateral, not visible from below yet visible in dorsal view, and not raised above head surface; iris operculum present. Six infraorbitals evident. Head plates and bones covered by short and flat odontodes. Dorsal surface of head with longitudinal keel on parieto-supraoccipital bone; compound pterotic ornamented with low reticulate ridges and pits. Mouth ovoid with lower lip larger than upper lip; ventral surface covered by wide oval papillae on upper lip and round papillae on lower lip; round papillae decreasing in size from oral aperture to lip margins; lip margins papillose. Platelets covering dorsal surface of upper lip. Each premaxilla with 13–25 teeth, each dentary with 13–22 teeth; premaxilla wider than dentary. Buccal papilla present, with papillose surface. Ventral surface of head covered by platelets without a particular arrangement. Maxillary barbel very short and projecting only slightly form mouth margin.
Abdomen flat, covered with two complete rows of plates and median incomplete row with two to four plates. Ventral plates angled and delimiting abdomen laterally. Posteriormost abdominal plate pair without median platelet. Anal plate arrow-head shaped with posterior margin either straight or slightly curved. Four lateral plate rows on body, all of them forming longitudinal keels.
Pectoral fin with distal margin straight, leading ray twice as thick as branched rays; median row of hypertrophied odontodes on dorsal surface absent, dorsal odontodes flat and not hypertrophied, ventral odontodes flat and medially directed. Pelvic fin with posterior margin slightly curved; dorsal odontodes flat, not hypertrophied, ventral odontodes flat, medially directed. Dorsal fin with posterior margin straight, triangular in overall shape. Anal fin the same as dorsal fin. Caudal fin emarginated with dorsal lobe larger than ventral lobe; dorsal filament present.
Coloration in alcohol. Head, dorsum, and caudal peduncle light brown; sides of head and body with longitudinal dark brown stripe continuous from tip of snout to fusion of median and ventromedian plate rows; ventral portions of head, body, and caudal peduncle light khaki from snout to base of caudal fin; some individuals show abdomen lighter than ventral portions of head, body, and caudal peduncle, so lateral darker stripe appears less evident from remaining coloration. Head with dark pigment delimiting plates in dorsal and lateral views; longitudinal dark brown stripe present from parieto-supraoccipital to dorsal-fin insertion. Dorsal, pectoral, pelvic, and anal fins with hyaline membranes and black spots on rays poorly arranged in bars. Caudal fin with overall dark brown-black coloration on both membranes and rays; area free of pigment present posteriorly at level of caudal emargination; tips unpigmented; variable clear ocelli present on dorsal and ventral lobes toward margins of lobes.
Ontogenetic variation. Description based on three paratype juveniles, 75.6–76.9 mm LS, and the adult male holotype, 112.3 mm SL. Cephalic ridges are more prominent in juveniles, in adults such structures become less defined and show gentle curvatures when compared with juveniles. Odontodes on plates on head, body and caudal peduncle form prominent keels no longer present in subadults and adults. Caudal is not visible in the holotype because of damage in the caudal fin, but present in juveniles. Spots on fins increase in number with size, from three to seven. Despite being less firm, odontodes on ventral surface of paired fins show the same median orientation as in adults.
Distribution. Farlowella yarigui is currently known only from the type locality in the middle río Magdalena Basin, western flank of the Cordillera Oriental, Colombia. The río Topón drains the western flank of the area under protection by the Parque Nacional Natural Serranía de los Yariguíes ( Fig. 2 View FIGURE 2 ).
Habitat. The type series of Farlowella yarigui was collected among wood sticks and partially-submerged vegetation using a bottom net. During a two-week collecting period, specimens of the new species were found exclusively in such microhabitat despite collection efforts throughout the main channel of the río Topón. Syntopic species included Astyanax caucanus , A. magdalenae , A. filiferus , Roeboides dayi , Sturisoma cf. panamense , Chaetostoma sp., Lasiancistrus caucanus , Creagrutus cf. guanes and Hemibrycon sp.
During the fieldtrip a wide range of microhabitats were sampled for fishes, but specimens of F. y a r i g u i were collected only with bottom netting amidst submerged vegetation and sticks ( Fig. 3 View FIGURE 3 ) in almost the same microhabitat where cis-Andean species of Farlowella are common. No specimens were detected in still water, mountain brooks, muddy environments, sandy banks, or among gravel and boulders in swift water.
It is noteworthy that such a distinctive loricariid catfish remained unnoticed until now in the río Magdalena Basin. Most of the collection efforts in Colombia have concentrated on this drainage (in addition to the Llanos region) since the XIX century. Some of the prominent collectors working in trans-Andean Colombia were Carl H. Eigenmann, Cecil Miles, George Dahl, Plutarco Cala, and Germán Galvis, along with several other incidental collectors. In addition, it was quite easy to secure specimens of F. y a r i g u i when working in the right microhabitat, which suggests that either collectors working in the río Magdalena Basin have been concentrating on mainstream and lowland areas and devoting little or no effort to piedmont areas, or that this drainage (one of the supposedly best-sampled in Colombia) is indeed still poorly known. In addition to the present species of Farlowella , Torres- Mejía et al. (2012) described Astyanacinus yariguies from the Serranía de los Yariguíes in Santander, suggesting that some tributaries of the río Magdalena Basin are still poorly known and that more intensive biodiversity inventories may enhance the discovery of additional new species.
The río Magdalena system supports ca. 80% of the total Colombian population (32 million people). This extreme anthropic impact has caused deforestation, an increase in erosion and sediment transport, floodplain draining, and fisheries collapse, that together severely degrade the environmental quality of habitats available for fish communities ( Galvis & Mojica, 2007). Contrary to the long-held opinion about the completeness of our knowledge on the río Magdalena fishes, urgent efforts are required on at least two fronts: (1) biodiversity inventories in fragile systems such as piedmont tributaries, and (2) more efficient conservation policies. Without such measures, the rate of habitat degradation will continue to rise while the rate of knowledge acquisition will decrease, and the río Magdalena biodiversity will be lost even before being adequately documented.
Etymology. The specific epithet is an arbitrary combination of letters in allusion to the Parque Nacional Natural Serranía de los Yariguíes , Departamento de Santander, Colombia, in recognition of its conservation efforts in this region of the Eastern Colombian Andes. It is used as a noun in apposition.
Remarks. The present contribution records for the first time the genus Farlowella from trans-Andean drainages exclusive of the Maracaibo Basin, where two other species have been recorded ( F. curtirostra and F. taphorni ). However, in his monograph on the fishes of Northern Colombia, Dahl (1971:81) comments on the type locality of F. gracilis and its confusing report for the Cauca valley. Additionally, in a footnote, the editor, probably Jorge Hernández-Camacho, a Colombian naturalist very interested in birds, may have confused the collector’s name based on his knowledge of ornithology, suggesting that such locality is in fact a mistyping of a locality in the río Cauca Basin: "The type locality of Farlowella gracilis corresponds to the río Cauquita, Cali, Departamento de Valle [del Cauca], small affluent on the left margin of the upper río Cauca. The manuscript labels for material collected there by Batty have allowed erroneous transcriptions (río "Cauqueta", río "Caquetá"), and therefore generates confusion with the río Caquetá or Yapuará, in the Amazon basin. If no label change ever occurred in fact, the type specimen of F. gracilis must be considered to be collected in the río Cauca basin, which requires confirmation through collection of additional specimens." (Editor footnote, our translation; Dahl, 1971:82).
Regan (1904:303) reported the collector of the holotype of F. gracilis to be Mr. M. D. Eder, and makes no mention of any collector and/or donor with last-name Batty, who was a well-known collector of birds. J. H. Batty collected birds in Western and Central Colombia during the XIX century, particularly in the Cauca Valley and adjoining mountains ( Chapman, 1912), and none of the localities reported by Chapman belong to the Amazonian versant of the Colombian Andes. In addition, there is no way to link the collections of J. H. Batty to M. D. Eder, so every data at hand suggest that the interpretation presented in Dahl (1971) has no merit.
The historical limits of the Departamento de Cauca in Colombia in the XIX century were markedly different from those currently in use, and included the upper río Caquetá, as well as from the Darién mountain range to the north and the río Mira to the south, including part or all of the actual departments of Cauca, Valle del Cauca, Chocó, Antioquia, Quindío, Caldas, Risaralda, and Nariño. Even today, the Departamento de Cauca includes the upper río Caquetá drainage. This historical background allows one to consider the possibility of reporting the río Caquetá as belonging to the Departamento de Cauca, and is even more likely to be confounded as synonymous with Cauca Valley, particularly in the XIX century.
Given the inconsistence in the collector’s information discussed in Dahl (1971), the lack of evidence linking Batty and Eder, and the historical geographic setting of Colombia in the XIX century, we conclude that earlier comments about the type locality of F. gracilis are unfounded. Finally, Retzer & Page (1996) reported F. gracilis as endemic to the río Caquetá. This implies that F. gracilis is not a trans-Andean species as suggested in earlier literature ( Dahl, 1971), and that F. yarigui is the only species known with certainty to dwell in trans-Andean rivers exclusive of the Maracaibo basin.

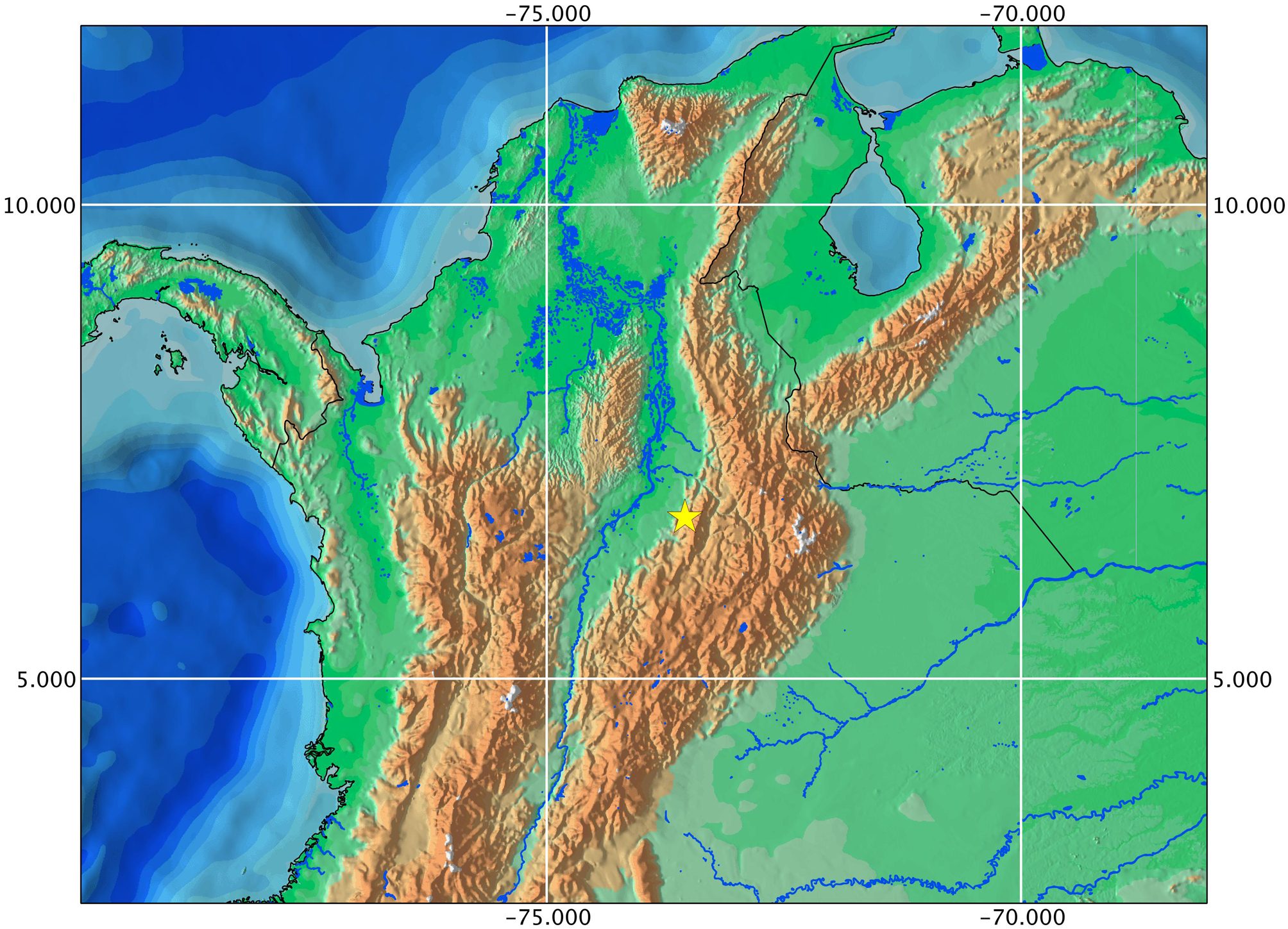

TABLE 1. Summary measurements for Farlowella yarigui n. sp. Measurements follow Boeseman (1976) with modifications. Measurements from head length to snout-mouth length are percentages of SL; and from eye diameter to head depth are percentages of HL. Measurements were taken from the holotype and the three paratypes (n = 4).
| Measurement | Range | Mean | SD |
|---|---|---|---|
| Standard length (SL) | 36.2–112.3 | 68.9 | 30.5 |
| Head length (HL) | 24.4–26.5 | 25 | 0.9 |
| Body depth at dorsal origin | 5.6–6.1 | 5.8 | 0.2 |
| Body width at dorsal origin | 5.3–6.8 | 5.9 | 0.6 |
| Body width at anal origin | 5.6–6.6 | 5.9 | 0.4 |
| Predorsal length | 42.1–47.0 | 44.9 | 1.9 |
| Postdorsal length | 48.0–52.6 | 50.1 | 2.0 |
| Postanal length | 48.8–52.1 | 50.1 | 1.4 |
| Caudal peduncle depth | 1.2–1.4 | 1.3 | 0.1 |
| Dorsal-spine length | 14.6–16.7 | 15.4 | 0.9 |
| Pectoral-spine length | 11.5–13.1 | 12.3 | 0.7 |
| Pelvic-spine length | 7.0–9.3 | 8.2 | 0.9 |
| Anal-spine length | 13.8–15.4 | 14.8 | 0.7 |
| Snout length | 15.0–16.4 | 15.9 | 0.7 |
| Snout-mouth length | 5.0–7.1 | 5.9 | 0.7 |
| Eye diameter | 8.4–10.9 | 9.5 | 1.1 |
| Interorbital width | 20.9–25.9 | 24.1 | 2.0 |
| Head width | 33.7–42.1 | 37.4 | 3.2 |
| Head depth | 21.7–27.8 | 24.7 | 2.5 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.