
Sauropodomorphs
|
publication ID |
https://doi.org/ 10.7717/peerj.4417 |
|
DOI |
https://doi.org/10.5281/zenodo.5614768 |
|
persistent identifier |
https://treatment.plazi.org/id/03FE9007-FFAA-831E-FE21-F8BD78D9D7EB |
|
treatment provided by |
Plazi |
|
scientific name |
Sauropodomorphs |
| status |
|
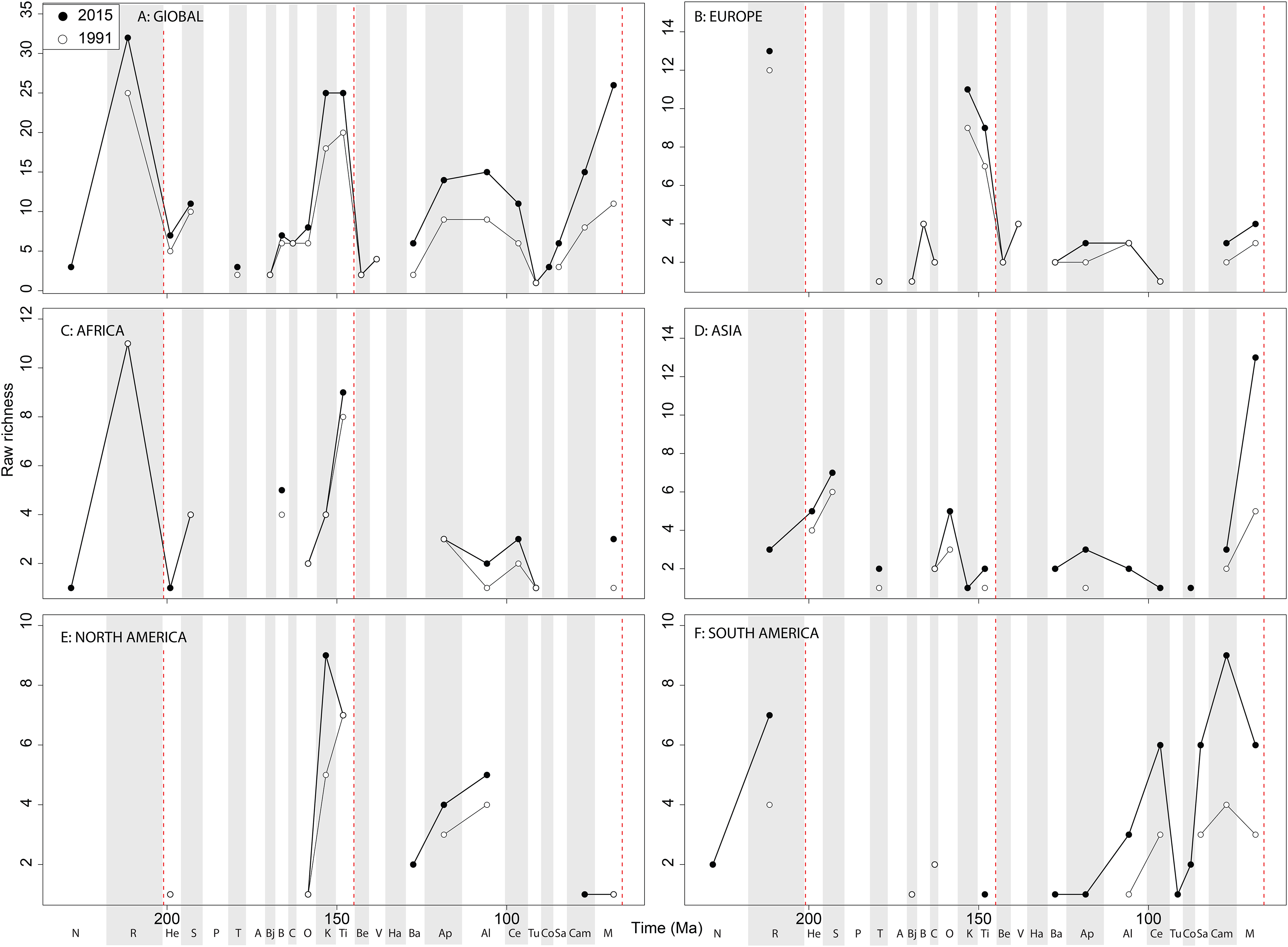
Sauropodomorph empirical diversity emphasises some more changes in raw patterns through publication time, particularly in the ‘middle’ and Late Cretaceous ( Fig. 11A View Figure 11 ).
Late Jurassic patterns are fairly consistent, with a rising Kimmeridgian and Tithonian raw diversity emphasising an apparent major decline across the J/K interval. In Europe, sauropods show a consistent and major decline in raw diversity from the Kimmeridgian to the Berriasian ( Fig. 11B View Figure 11 ). Much of the rest of the Cretaceous is too poorly sampled, but raw sauropod diversity never attains Kimmeridgian levels in Europe for the rest of their evolutionary history. Sauropodomorph dinosaurs are generally better sampled than theropods and ornithischians in Africa, showing an apparent decline through the Triassic/ Jurassic transition, a latest Jurassic raw diversity peak, and low levels through the ‘middle’ to Late Cretaceous transition ( Fig. 11C View Figure 11 ). In Asia, raw taxonomic diversity is generally low compared to the Maastrichtian, in which diversity is relatively high and still rapidly increasing through publication history ( Fig. 11D View Figure 11 ). The North American sauropod record is very patchy, with the latest Jurassic showing a shift from rapidly increasing raw diversity from the Oxfordian to a slight drop from the Kimmeridgian to Tithonian ( Fig. 11E View Figure 11 ).
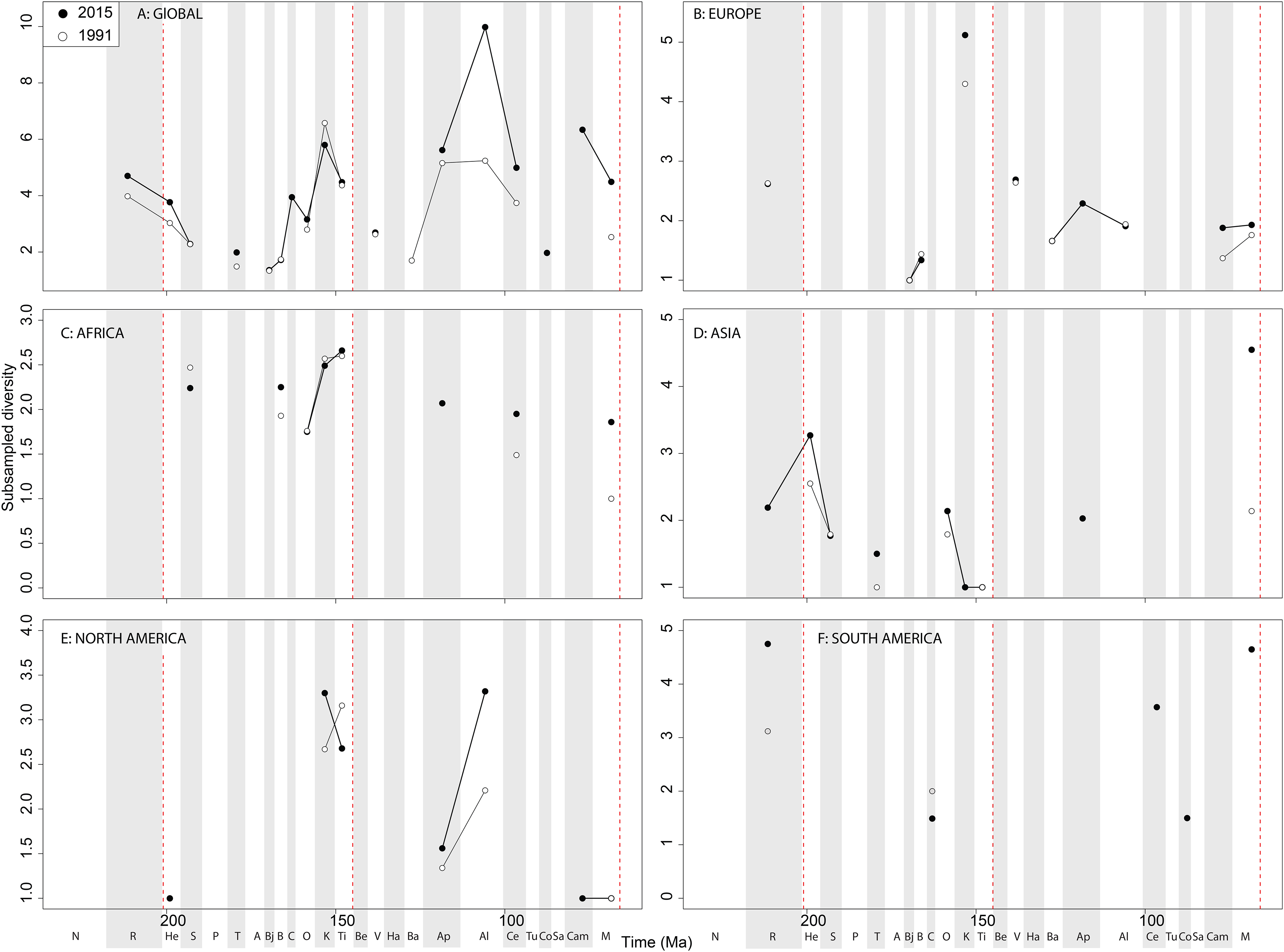
The South American Jurassic sauropod record is patchy, but raw diversity is increasing throughout the ‘middle’ to Late Cretaceous through publication history ( Fig. 11F View Figure 11 ). At a ‘global’ level, Jurassic sauropodomorph subsampled diversity remains consistent through publication history ( Fig. 12A View Figure 12 ). Here, we see steadily increasing diversity levels through the Middle and Late Jurassic, before a decline through the J/K transition, which might have been initiated before the J/K boundary itself. The greatest change in subsampled diversity is in the Albian, which has almost doubled in the last 20 years, with implication for the ‘mid-Cretaceous sauropod hiatus’ ( Mannion & Upchurch, 2011). Subsampling reduces the European diversity signal due to poor sampling of sauropods, although there is evidence for the sauropod decline beginning prior to the J/K transition ( Fig. 12B View Figure 12 ). In Africa, when subsampling is applied, the few intervals in which a signal emerges reveal a fairly constant level of diversity through the Jurassic and Cretaceous, and through publication time, with the notable exception being an increase in subsampled diversity in the latest Jurassic ( Fig. 12C View Figure 12 ). In Asia, the signal is also fairly poor after subsampling is applied ( Fig. 12D View Figure 12 ). Here, we see an increase in subsampled diversity across the Triassic/Jurassic transition, and the highest diversity level is in the Maastrichtian, where subsampled estimates have increased by more than double in the last 20 years. In North America, the subsampled signal is highly degraded, although of note is a near doubling of Albian diversity levels in the last 20 years ( Fig. 12E View Figure 12 ). In South America, the signal is very inconsistent, but improving through publication history, with a patchy Late Cretaceous signal beginning to emerge ( Fig. 12F View Figure 12 ). Full subsampling results are provided in Supplemental Informations 7 and 8.
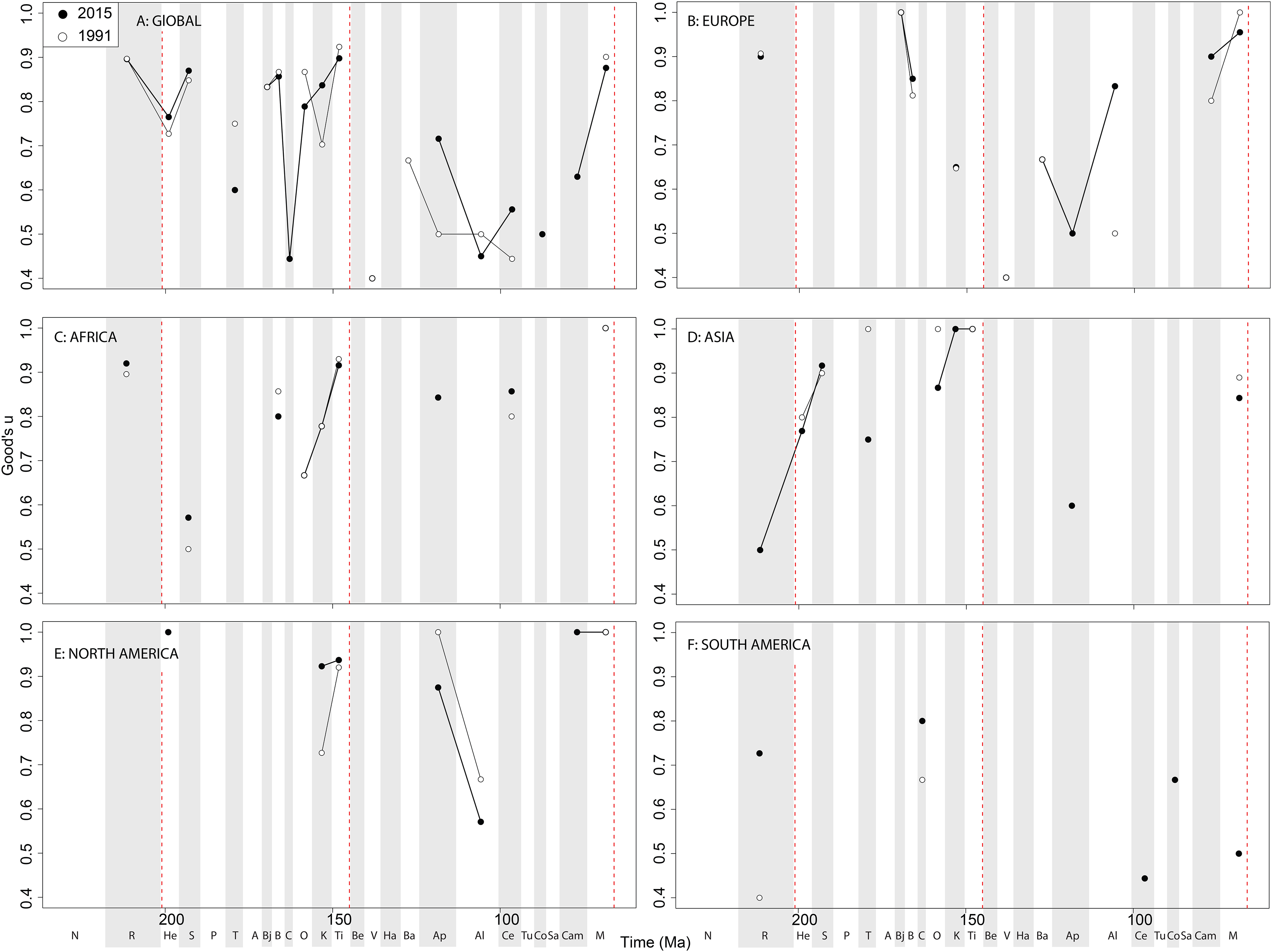
Sauropodomorph coverage varies greatly at the ‘global’ level, with high levels in the Triassic-Jurassic transition, the Middle and Late Jurassic (with the exception of the Callovian), and the Maastrichtian ( Fig. 13A View Figure 13 ). As with theropods and ornithischians, however, this is a composite of a very patchy regional record. In Europe, coverage is high during the latest Triassic, Middle Jurassic, and Late Cretaceous, and this does not seem to have varied with publication time ( Fig. 13B View Figure 13 ). In Africa, moderate levels of coverage also have not changed substantially since 1991 ( Fig. 13C View Figure 13 ). In Asia, coverage is generally high in the Late Jurassic, but the Cretaceous record is incredibly poor with just two data points (Aptian and Maastrichtian; Fig. 13D View Figure 13 ). In North America, the latest Jurassic has high coverage levels, which are increasing through publication history in the Kimmeridgian, and moderately high coverage in the Aptian and latest Cretaceous ( Fig. 13E View Figure 13 ). In South America, coverage is very patchy and inconsistent, with the only noteworthy change through publication history being an increase for the Rhaetian interval ( Fig. 13F View Figure 13 ).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.