
Fabrosaurus australis, Ginsburg, 1964
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2005.00182.x |
|
persistent identifier |
https://treatment.plazi.org/id/03FE87B6-124E-6047-FF58-9B18FD15C967 |
|
treatment provided by |
Diego |
|
scientific name |
Fabrosaurus australis |
| status |
|
FABROSAURUS AUSTRALIS GINSBURG, 1964
Diagnosis: Fabrosaurus australis is not diagnosable on the basis of unique characters, or on the basis of a unique combination of characters, and is here regarded as a nomen dubium; the holotype is considered Ornithischia indet.
Holotype: MNHN LES9, partial dentary with three teeth ( Ginsburg, 1964; Galton, 1978: fig. 6; Sereno, 1991: fig. 1; Thulborn, 1992: fig. 1).
Horizon: Upper Elliot Formation (Hettangian– Sinemurian), Likhoele, Lesotho.
Discussion: There has been substantial debate regarding whether the holotype of Fabrosaurus (MNHN LES9) possesses any unique characters that would support the validity of the genus. Charig & Crompton (1974: 167–168) argued that the teeth of Fabrosaurus were symplesiomorphic for Ornithischia , and therefore undiagnostic; they considered it a nomen dubium. Galton (1978) considered Fabrosaurus as valid, but removed the specimens referred to Fabrosaurus by Thulborn (1970, 1971a, 1972) into a new genus, Lesothosaurus diagnosticus . He suggested that Fabrosaurus was distinct on the basis of its broader dentary, possession of ‘special foramina’, and minor differences in tooth height/width ratios. Gow (1981) showed that all of these features were undiagnostic, and could be the result of ontogenetic or individual variation. Sereno (1991) redescribed the holotype of Fabrosaurus ( Sereno, 1991: fig. 1); he concluded that the holotype lacked autapomorphies and could not be distinguished from other basal ornithischians.
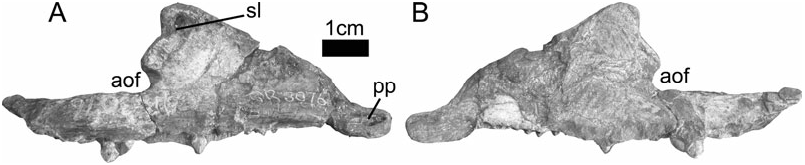
Thulborn (1992: 286) and Hunt & Lucas (1994: 238) have argued that no other ornithischian taxon shares the same tooth morphology as Fabrosaurus and that, although autapomorphic characters are clearly absent, the holotype dentary can be diagnosed by a unique combination of primitive characters [i.e. a metataxon (sensu Gauthier, 1986)]. Knoll (2002b: 238) noted that the same tooth morphology is present in an undescribed basal ornithischian from the Upper Elliot Formation; this demonstrates that the tooth morphology of Fabrosaurus is shared by more than one taxon, with the consequence that Fabrosaurus should be considered a nomen dubium. The taxonomic affinities of the specimen (NM QR 3076) used by Knoll to support this supposition are here considered uncertain (see below). However, the same tooth morphology is also recognized in an undescribed new taxon from the Lower Elliot Formation (SAM-PK-K8025) and is very similar to that of other basal ornithischians such as Scutellosaurus lawleri (see Colbert, 1981: figs 9, 10; Sereno, 1991: 173–174). This tooth morphology is therefore widespread in basal ornithischians, as first suggested by Charig & Crompton (1974: 167–168). Here I follow most recent authors ( Weishampel & Witmer, 1990a; Sereno, 1991; Knoll, 2002a, b; Norman, Witmer & Weishampel, 2004a) in considering Fabrosaurus australis to be a nomen dubium. Lesothosaurus diagnosticus is accepted as the valid binomial for the specimens referred to ‘ Fabrosaurus ’ by Thulborn (1970, 1971a, 1972). Although the holotype of ‘ Fabrosaurus ’ (MNHN LES9) is clearly ornithischian, its phylogenetic position within the clade cannot be determined owing to the fragmentary nature of the specimen and its lack of phylogenetically informative character states; for this reason it should be considered as Ornithischia indet. ( Sereno, 1991).
LESOTHOSAURUS DIAGNOSTICUS GALTON, 1978
( FIGS 3A–B, E–F View Figure 3 , 4 View Figure 4 , 24B View Figure 24 )
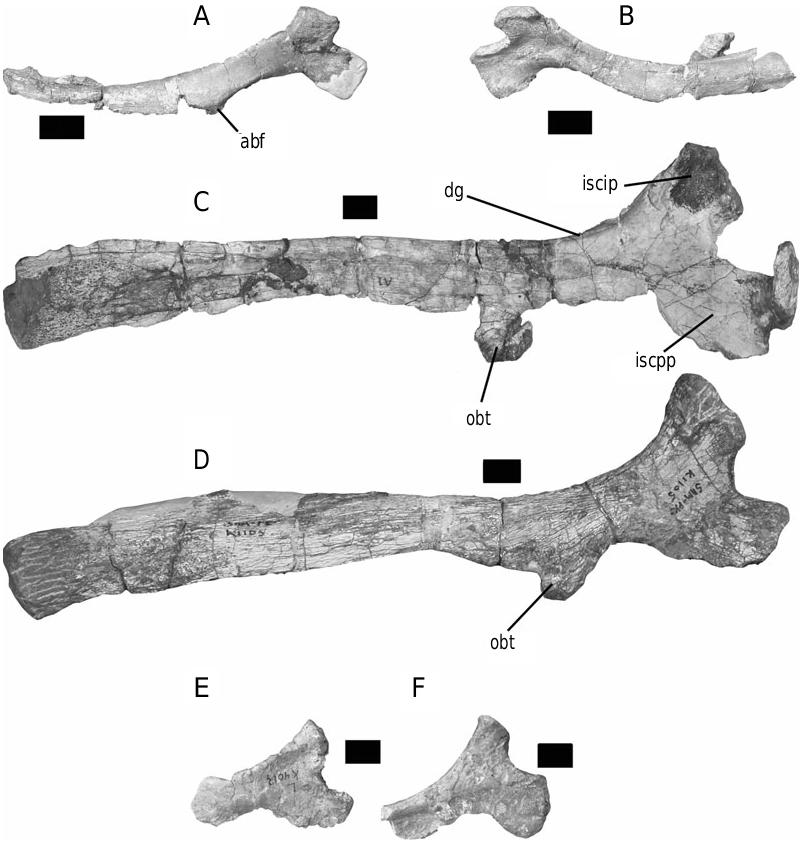
Diagnosis: Lesothosaurus diagnosticus cannot be diagnosed on the basis of autapomorphic features. However, it is distinguishable from all other ornithischians on the basis of a unique combination of primitive and derived character states and should be considered a metataxon (sensu Gauthier, 1986), pending further discoveries. This unique character combination comprises: anterior premaxillary foramen present; slot in maxilla for lacrimal present; six premaxillary teeth present; absence of diastema between the premaxillary and maxillary teeth; maxillary teeth lack apicobasally extending ridges on their lingual and labial faces; manual phalanges lacking prominent intercondylar processes; ilium with welldeveloped supraacetabular flange and ventromedially angling brevis shelf visible in lateral view; dorsal groove on the ischial shaft present; shaft of ischium twists through 90° along its length, forms an elongate symphysis with the opposing ischial blade, and lacks a tab-shaped obturator process; prepubic process short and mediolaterally flattened rather than rodlike and does not extend beyond the end of the preacetabular process of the ilium; postcranial osteoderms absent.
Syntypes: BMNH RUB17, mostly disarticulated remains of at least two individuals, one larger than the other, including most of one articulated skull ( Figs 3A–B View Figure 3 , 4 View Figure 4 , 24B View Figure 24 ; Thulborn, 1970: figs 2–7; Thulborn, 1971a: figs 1–4, 8; Thulborn, 1972: figs 2–12; Galton, 1978: fig. 2; Sereno, 1991: figs 6, 7, 8B–F, 9A– B, E–F); BMNH RUB 23, partial skull ( Thulborn, 1970: fig. 1; Thulborn, 1971a: fig. 7).
Referred material: BMNH R11956, partial skull; BMNH R8501, nearly complete, disarticulated skull ( Sereno, 1991: fig. 2); BMNH R11004, partially articulated posterior skull and anterior neck, including the braincase, parietals, right squamosal, right quadrate, right posterior lower jaw, axis and third cervical ( Sereno, 1991: fig. 8A; SAM-PK-K400, partial postcranium ( Santa Luca, 1984: figs 10, 11); SAM-PK-K401, partial postcranium, including proximal ischia ( Fig. 3E–F View Figure 3 ; Santa Luca, 1984: figs 12, 13, 18, 20, 24, 25, 27–32); SAM-PK-K1106, partial postcranium, including proximal ischia ( Santa Luca, 1984: figs 1–9, 19, 21–23, 26).
Horizon: All known specimens of Lesothosaurus are derived from the Upper Elliot Formation (Lower Jurassic: Hettangian-Sinemurian) of South Africa and Lesotho. Locality details can be found in Thulborn (1970, 1972) and Santa Luca (1984).
Discussion: Sereno (1991: 172) revised the diagnosis of Lesothosaurus and suggested six characters as potential autapomorphies, but noted that: ‘they may also characterize an undescribed larger ornithischian from the same formation’. Subsequent work and description of new taxa has shown that most of these characters have a more widespread distribution than realized previously ( Peng, 1992, 1997; Xu, Wang & You, 2000), and most of them appear to be present in the larger basal ornithischian (described below as Stormbergia dangershoeki gen. et sp. nov.) from the same formation. The distribution of the six diagnostic features of Lesothosaurus proposed by Sereno (1991) is discussed below:
1. ‘Slot in maxilla for lacrimal’. Peng (1997) reported the presence of this feature in Agilisaurus louderbacki Peng, 1990; a basal ornithischian from the Middle Jurassic of Sichuan, China. It has also been reported for Jeholosaurus shangyuanensis Xu et al., 2000 , from the Early Cretaceous of Liaoning ( Xu et al., 2000: 320). However, the holotype skulls of both Agilisaurus (ZDM T6011), and Jeholosaurus (IVPP V12529 View Materials ), are preserved in articulation and the nature of the maxilla-lacrimal contact cannot be confirmed (pers. observ.). Knoll (2002a, b) reports the presence of this feature in NM QR 3076, a specimen which he considers to represent a ‘large fabrosaurid’. Although the feature does indeed appear to be present ( Fig. 2 View Figure 2 : sl), NM QR 3076 is here considered to be of uncertain affinities (see below). The distribution of this character must thus be considered uncertain at present, and so cannot be confirmed as autapomorphic for Lesothosaurus .
2. ‘Anterior premaxillary foramen’. Most ornithischians have a premaxillary foramen positioned at the anteroventral corner of the external naris. In Lesothosaurus (BMNH RUB17, R8501) a groove passes anteroventrally from the premaxillary foramen to a second foramen which Sereno (1991) termed the anterior premaxillary foramen. This foramen opens into a canal that appears to open onto the premaxillary palate just anterior to the first premaxillary tooth ( Sereno, 1991). Knoll (2002a, 2002b) notes that the feature is present in NM QR 3076, but this specimen is here considered to be of uncertain affinities (see below). An anterior premaxillary foramen in an equivalent position is present on the lateral margin of the premaxilla of Hypsilophodon foxii, Huxley, 1869 (BMNH R2477) and is connected to the premaxillary foramen by a shallow incised groove. The presence of this foramen was noted by Sereno (1991) who claimed that it opened into an internal cavity of the premaxilla rather than onto the premaxillary palate. However, the anteriormost portion of the premaxilla of BMNH R2477 is damaged and it is not possible to ascertain the position of the canal opening. It seems possible that the anterior premaxillary foramen of Hypsilophodon is homologous with that of Lesothosaurus . The premaxillae of many basal ornithischians are unknown (e.g. Bonaparte, 1976), or poorly preserved anteriorly (e.g. Colbert, 1981; Haubold, 1991), so this character may have had a wider distribution than previously realized. Although this feature may turn out to be an autapomorphy of Lesothosaurus , this cannot be confirmed at present.
3. ‘Short forelimb’. The forelimb is highly reduced in Agilisaurus (ZDM T6011; Peng, 1992, 1997), Stormbergia (SAM-PK-K1105; Table 2) and in an undescribed basal ornithischian from the Upper Triassic Lower Elliot Formation of South Africa (SAM-PK-K8025). This character cannot be considered autapomorphic for Lesothosaurus .
4. ‘Lateral exposure of brevis surface on the postacetabular process [of the ilium]’. As noted by Sereno (1991: 172) this feature is present in Scelidosaurus (BMNH R1111, R6704, see Fig. 24B View Figure 24 ). It is also seen in Agilisaurus ( Peng, 1992: fig. 5; Peng, 1997; ZDM T6011), Stormbergia (BMNH R11000, see Fig. 13 View Figure 13 ) and in the undescribed Lower Elliot Formation taxon (SAM-PK-K8025). Indeed, this feature appears to be plesiomorphic for Ornithischia as a whole ( Knoll, 2002a, b).
5. ‘Dorsal groove on the ischial shaft’. This feature is present in Agilisaurus ( Peng, 1997; ZDM T6011),
in the undescribed Lower Elliot Formation taxon (SAM-PK-K8025), and weakly present in some individuals of Stormbergia (BMNH R11000, Fig. 3C View Figure 3 ; Knoll 2002a, b), although apparently not in others (SAM-PK-K1105, Fig. 3D View Figure 3 ). Therefore, this feature cannot be considered autapomorphic for Lesothosaurus diagnosticus . 6. ‘Pedal digit 1 reduced’. The reduction of pedal digit 1 occurs widely in basal ornithischians including Agilisaurus ( Peng, 1992: fig. 6C; ZDM T6011), Othnielia rex Galton, 1977 ( Galton & Jensen, 1973: fig. 6D, ‘ Yandusaurus’ multidens ( He & Cai, 1984: fig. 20C; ZDM T6001) and Jeholosaurus ( Xu et al., 2000; IVPP V12529 View Materials ) and also occurs in Stormbergia (SAM-PK-K1105, see Fig. 17B View Figure 17 ). This feature cannot be considered autapomorphic for Lesothosaurus diagnosticus .
In addition, Norman et al. (2004a) suggest that Lesothosaurus is characterized by: a prominent crested obturator process on the ischium, and metatarsal V was either absent or ‘strongly reduced’. However, as argued below, a true tab-shaped obturator process is absent in Lesothosaurus (BMNH RUB17, Figs 3A–B View Figure 3 , 4 View Figure 4 ). The distinct ventral corner that occurs in a similar position on the ischium is not unique to Lesothosaurus , but is present in the undescribed Lower Elliot Formation ornithischian (SAM-PK- K8025) and may be present in the basal thyreophoran Scutellosaurus lawleri (UCMP 130580: Rosenbaum & Padian, 2000: figs 2, 4) and the basal genasaurian Agilisaurus (ZDM T6011; pers. observ.). The apparent complete loss of metatarsal V is also seen in Stormbergia (SAM-PK-K1105, see Fig. 17B View Figure 17 ). In many other basal ornithischians such as Agilisaurus ( Peng, 1992: fig. 6) and Othnielia ( Galton & Jensen, 1973: fig. 6A) metatarsal V is extremely reduced.
Consequently, Lesothosaurus diagnosticus is difficult to diagnose on the basis of autapomorphies. However, the syntype specimens BMNH RUB17 and BMNH RUB23 can be differentiated from all other known basal ornithischians; including: Pisanosaurus mertii Casamiquela, 1967 from the Late Triassic of Argentina ( Bonaparte, 1976; Sereno, 1991); an undescribed basal ornithischian from the Late Triassic of South Africa (SAM-PK-K8025); Stormbergia dangerhoeki gen. et sp. nov. and the heterodontosaurids Heterodontosaurus tucki ( Santa Luca, 1980) and Abrictosaurus consors ( Thulborn, 1974) , all of which are from the Lower Jurassic of South Africa; the basal thyreophoran Scutellosaurus lawleri from the Lower Jurassic of Arizona ( Colbert, 1981; Rosenbaum & Padian, 2000); and Agilisaurus louderbacki and ‘ Yandusaurus’ multidens, both from the Middle Jurassic of China ( He & Cai, 1984; Peng, 1992, 1997).
Pisanosaurus mertii is known from a single fragmentary skeleton, collected from the Ischigualasto Formation (Carnian) of Argentina, and has been considered the basalmost known ornithischian ( Sereno, 1991). There is some doubt as to whether the elements which comprise the partial skeleton belong together, or whether the taxon is a composite ( Sereno, 1991). Nearly all elements show substantial differences from Lesothosaurus . The preserved cranial elements demonstrate features apparently derived with respect to Lesothosaurus , in particular the presence of welldeveloped wear facets on the teeth. Postcranially, however, Pisanosaurus appears very primitive, possibly lacking a prepubic process, and with a poorly developed lateral expansion of the distal tibia ( Bonaparte, 1976; Sereno, 1991).
A single ornithischian specimen (SAM-PK-K8025) known from the Late Triassic Lower Elliot Formation of South Africa appears to represent a new taxon of basal ornithischian (R. J. Butler & R. M. H. Smith, unpubl. data) and can be distinguished from Lesothosaurus by several features, including the retention of a relatively large manus, the phalanges of which have prominent proximal intercondylar processes and distinct dorsal pits on their distal ends. These features are absent in Lesothosaurus (BMNH RUB17; Sereno, 1991, fig. 8B).
The anatomical features which distinguish Lesothosaurus from Stormbergia dangerhoeki gen. et sp. nov. are discussed below.
Heterodontosaurus tucki , Abrictosaurus consors , Echinodon becklesii ( Norman & Barrett, 2002) and undescribed taxa from the Lower Jurassic Kayenta Formation of Arizona (P. C. Sereno, pers. comm., 2004) and the Upper Jurassic Morrison Formation of Colorado (P. M. Galton, pers. comm., 2004), apparently form a clade of heterodontosaurids ( Weishampel & Witmer, 1990b; R. J. Butler, unpubl. data), which can be distinguished from Lesothosaurus by a large number of derived features. These include: premaxillary tooth count reduced to three or less, arched diastema present between premaxilla and maxilla, wedgeshaped predentary present, supraacetabular flange of the ilium absent, brevis shelf reduced and horizontal.
Scutellosaurus lawleri , a primitive thyreophoran known from numerous partial skeletons ( Colbert, 1981; Rosenbaum & Padian, 2000; MCZ unpubl. material) from the Kayenta Formation of Arizona, is plesiomorphic in most of its features, and no autapomorphies appear to be present (although it can be diagnosed using a combination of primitive and derived characters). Its anatomy is therefore very similar to that of Lesothosaurus , but it can clearly be distinguished from the latter by the presence of postcranial osteoderms. As yet, no ornithischian osteoderms have been found in the Elliot Formation.
Agilisaurus louderbacki is known from an articulated and nearly complete skeleton (ZDM T6011) from the Lower Shaximiao Formation (Middle Jurassic) of Dashanpu, Sichuan Province, China. Although Agilisaurus has been considered by some authors as a basal ornithopod ( Sues & Norman, 1990; Weishampel & Heinrich, 1992; Weishampel et al., 2003), it is apparently a more basal ornithischian (Barrett, Butler & Knoll, in press) with numerous similarities (e.g. the presence of a groove on the ischium, shortened forelimb and reduced pedal digit one) to Lesothosaurus ( Peng, 1997) . However, these similarities are probably plesiomorphic as Agilisaurus has a large number of derived features absent in Lesothosaurus . These include: premaxillary tooth count reduced to five, prepubic process elongated, reaching almost to the end of the preacetabular process of the ilium (ZDM T6011; Peng, 1997), and a tab-shaped obturator process on the ischium ( Peng, 1992, 1997). Agilisaurus additionally has a number of unusual, autapomorphic, features which are absent in Lesothosaurus (pers. observ. of ZDM T6011; Barrett et al., in press), including the presence of a strongly heterodont dentary dentition and a palpebral bone which traverses the entire width of the orbit.
The alpha-level taxonomy of the genus Yandusaurus is currently unstable. Barrett et al. (in press) consider the species ‘ Yandusaurus’ multidens to be generically distinct from the genus Yandusaurus and the genus Agilisaurus. This conclusion is followed herein, with ‘ Yandusaurus’ multidens being represented by the holotype (ZDM T6001) and paratype (ZDM T6002) specimens, which are both from the same locality as the holotype of Agilisaurus louderbacki (see above). ‘ Y ’ multidens retains numerous ornithischian plesiomorphies (ZDM T6001; He & Cai, 1984) but has a large number of derived features which clearly distinguish it from Lesothosaurus , including the presence of some apico-basally extending ridges on the maxillary crowns (ZDM T6001), the absence of a supraacetabular flange on the ilium, a horizontal, rather than ventromedially angling, brevis shelf (ZDM T6001, pers. observ.), and a prepubic process which is elongate and rod-shaped, and extends beyond the anterior end of the preacetabular process ( He & Cai, 1984: fig. 17A).
STORMBERGIA DANGERSHOEKI GEN. ET SP. NOV.
( FIGS 3C–D View Figure 3 , 5 View Figure 5 , 7–19 View Figure 7 View Figure 8 View Figure 9 View Figure 10 View Figure 11 View Figure 12 View Figure 13 View Figure 14 View Figure 15 View Figure 16 View Figure 17 View Figure 18 View Figure 19 )
Etymology: Genus name refers to the Stormberg Group of South Africa and Lesotho, the rock sequence that has provided so much information on early dinosaurs. Species epithet refers to the locality from which the holotype (SAM-PK-K1105) was collected.
Diagnosis: A relatively large (around 2 m length) basal ornithischian, lacking unambiguous autapomorphies but distinguished by a unique combination of character states, not seen in other basal ornithischians, including: ilium with a robust, elongate, pubic peduncle, well-developed supra-acetabular flange and ventromedially angling brevis shelf; acetabulum partially closed medially by a ventral flange of the ilium; ischial symphysis present distally only; ischium with an essentially untwisted shaft and a well-developed, proximal tab-shaped obturator process; prepubic process short and mediolaterally flattened.
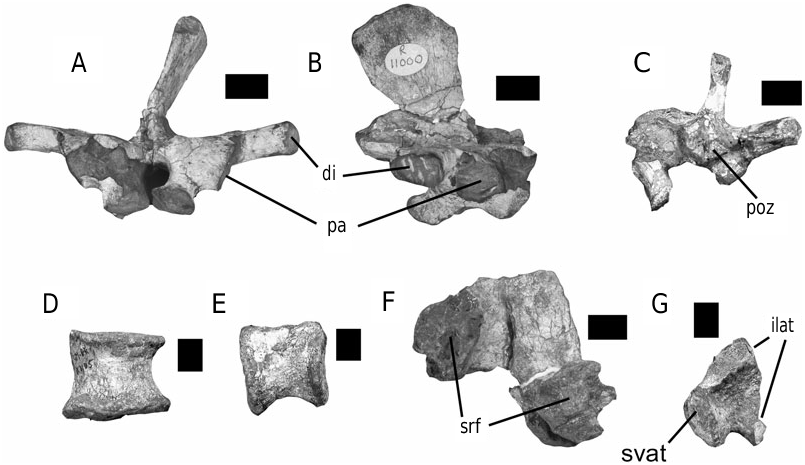
Holotype: SAM-PK-K1105 ( Figs 3D View Figure 3 , 7 View Figure 7 , 8C–G View Figure 8 , 9C–D View Figure 9 , 11 View Figure 11 , 12 View Figure 12 , 14–17 View Figure 14 View Figure 15 View Figure 16 View Figure 17 , 19 View Figure 19 ), a partial postcranial skeleton, including the axial neural arch, cervical neural arches and centra, dorsal neural arches and centra, two sacral neural arches and sacral ribs, caudal vertebrae, coracoids, fragmentary humeri, left radius and ulna, left ischium, proximal right ischium, pubes, femora, tibiae, tarsals and metatarsals, numerous disarticulated phalangeal elements. Collected from Dangershoek Farm, Herschel District, eastern Cape Province, Republic of South Africa by C. E. Gow. This specimen was previously on loan to the Museum of Comparative Zoology, Harvard, and has been referred to by Knoll (2002a: 601; 2002b: 242).
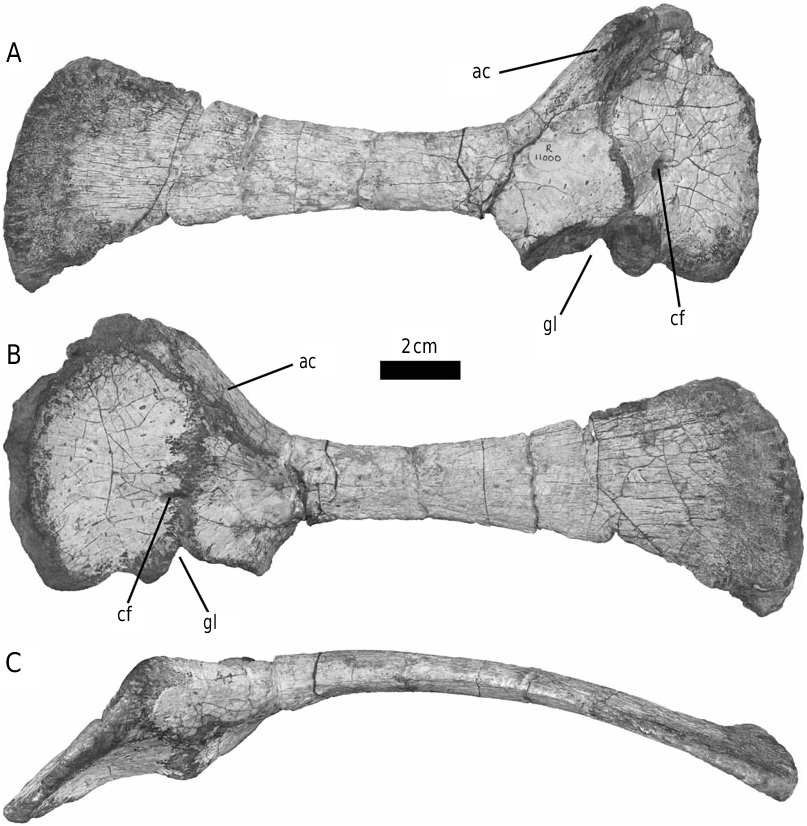
Paratype: BMNH R11000 ( Figs 3C View Figure 3 , 8A–B View Figure 8 , 9A–B View Figure 9 , 10 View Figure 10 , 13 View Figure 13 , 18 View Figure 18 ), partial postcranial skeleton, including a dorsal neural arch, complete proximal caudal vertebra, right scapulocoracoid, left ilium, right ischium, proximal left femur, distal left tibia and partial right fibula. This specimen was collected from Pokane, Lesotho by a 1966–67 British Museum/University of London expedition team, and has been referred to by Knoll (2002a: 601; 2002b: 242).
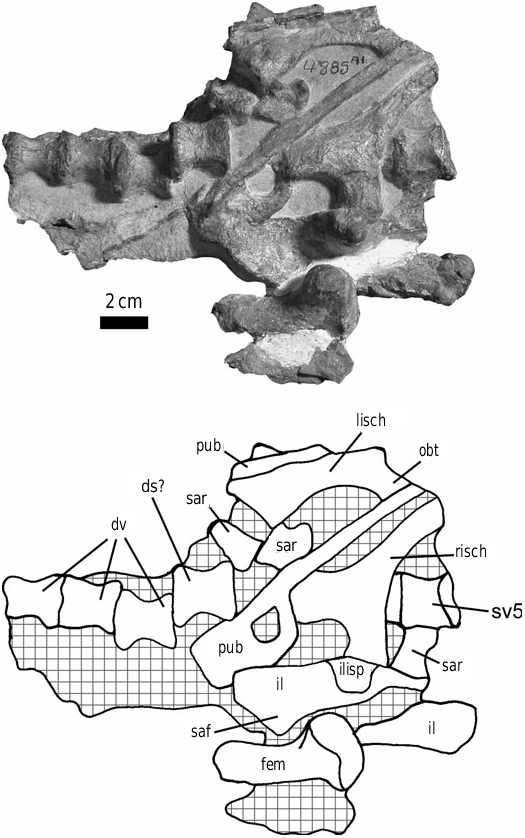
Referred specimen: BP/1/4885 ( Fig. 5 View Figure 5 ), in part, collected from Mequatling, Clocolon District, Free State Province by L. Huma in 1980. This specimen number includes portions of several different taxa (including the synapsid Tritylodon ), but an articulated portion is here considered to represent a juvenile individual of Stormbergia and was referred to by Knoll (2002a: 601; 2002b: 242) as an individual of Lesothosaurus diagnosticus .
Horizon: All known specimens are from the ‘Red Beds’ of the Upper Elliot Formation (Lower Jurassic: Hettangian–Sinemurian).
Discussion: As noted by Knoll (2002a: 601), the specimens here referred to Stormbergia are very similar in morphology to the syntype (BMNH RUB17) and referred material of Lesothosaurus diagnosticus . Indeed, Knoll (2002a) has speculated that Stormbergia may represent adults of Lesothosaurus diagnosticus , implying that the Lesothosaurus syntypes and referred material represent juveniles. The subadult status of the Lesothosaurus material may be supported by the lack of neurocentral fusion in the vertebral column and lack of sacral fusion (however, fusion is also absent in the individuals referred here to Stormbergia and may be a persistent feature of basal ornithischians).
sv5
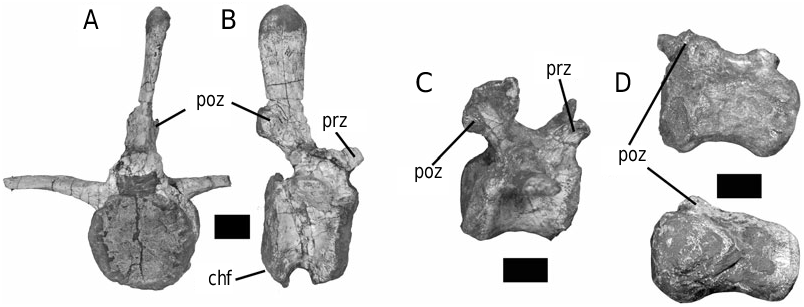
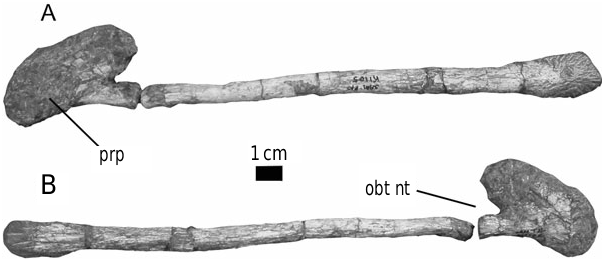
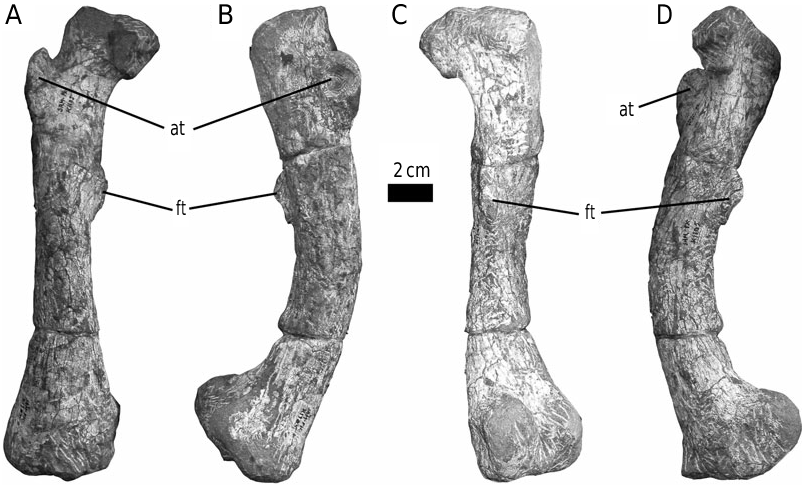
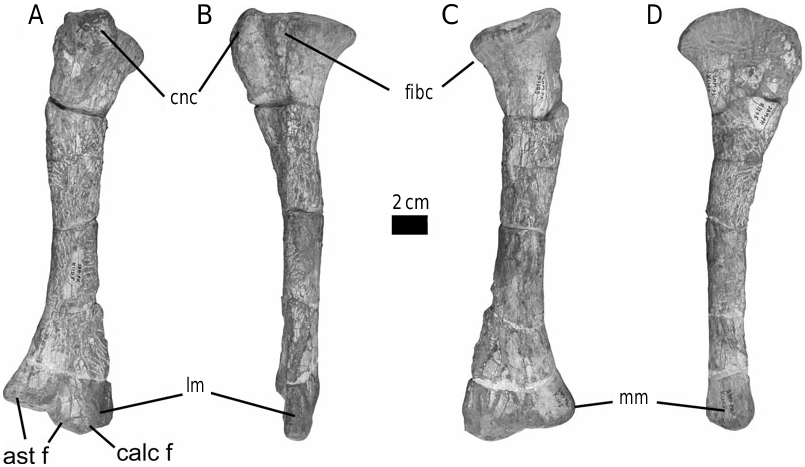
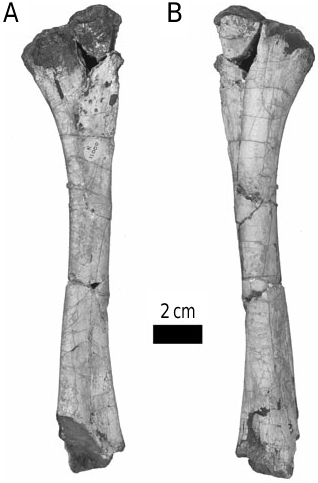
The form of the ischium clearly distinguishes Stormbergia from Lesothosaurus ( Figs 3–5 View Figure 3 View Figure 4 View Figure 5 ). The ischium of Stormbergia has a very well-defined, tabshaped obturator process (BMNH R11000, Fig. 3C View Figure 3 ; SAM-PK-K1105, Fig. 3D View Figure 3 ; BP/1/4885, Fig. 5 View Figure 5 ), similar to that seen in more derived ornithischians traditionally referred to Ornithopoda (e.g. Galton, 1974a). The presence or absence of an obturator process in Lesothosaurus has been much debated ( Sereno, 1986, 1991; Thulborn, 1992; Knoll, 2002a, b). In an undescribed new ornithischian from the Lower Elliot Formation (SAM-PK-K8025) the well-preserved proximal ischia show that a tab-shaped obturator process was definitely absent. Rather, the shaft is strongly twisted such that the medial surface turns to face dorsally, and its ventral or anterior surface (which actually is directed medially due to the torsion of the shaft) curves to form a distinct corner. The ischia of Lesothosaurus (BMNH RUB17, Figs 3A–B View Figure 3 , 4 View Figure 4 ) are very similar, also showing this strong torsion, the distinct corner (‘prominent angle’ of Sereno, 1991) and lacking development of a tab-shaped obturator process ( Sereno, 1991; pers. observ.). The suggestion that an obturator process was present in Lesothosaurus ( Thulborn, 1972) is based upon the left ischium of BMNH RUB17 ( Figs 3A View Figure 3 , 4 View Figure 4 ), which is damaged along its ventral surface. As noted by Sereno (1991), two fragments glued to this damaged ventral margin have been misidentified as the obturator process (see: Fig. 3A View Figure 3 ). The more complete right ischium clearly lacks this ‘process’ ( Fig. 3B View Figure 3 ; Sereno, 1991).
The strong torsion of the ischial shaft and the development of a distinct anteroventral corner to the shaft appear to be linked to the presence of an elongate ischial symphysis in Lesothosaurus ( Fig. 4 View Figure 4 ). The anteroventral corner marks the most proximal extension of this symphysis, which would have extended for at least 50% of the length of the ischium ( Fig. 4 View Figure 4 ). Sereno (1986) suggested that an ischial symphysis restricted to the distal end of the ischia was a synapomorphy of ornithischians; however, an elongate ischial symphysis appears to be retained in a number of basal ornithischians, including Lesothosaurus , an undescribed basal ornithischian from the Lower Elliot Formation (SAM-PK-K8025), Agilisaurus louderbacki (ZDM T6011; pers. observ.) and possibly Scutellosaurus lawleri (UCMP 130580: Rosenbaum & Padian, 2000: figs 2, 4). By contrast, the ischial symphysis of Stormbergia (SAM-PK-K1105, BMNH R11000) was present distally only, and the ischial shaft undergoes little torsion along its length. The loss of an elongate ischial symphysis is a derived state shared with most other ornithischians.
None of the specimens (SAM-PK-K401, Fig. 3E–F View Figure 3 ; SAM-PK-K1106) described by Santa Luca (1984: figs 18, 19) have the distal portions of their ischia preserved or exposed, and so do not demonstrate a tabshaped obturator process, but appear instead to be very similar in morphology to the ischia of BMNH RUB17 (contra Norman et al., 2004a: 331).
Knoll (2002b) has suggested that ‘the specimen BPI 4885 [sic] pleads for the presence of an obturator process in Lesothosaurus diagnosticus ’. This specimen number (correctly BP/1/4885) includes a large amount of material, much unprepared, which includes some definite ornithischian material, plentiful material of the synapsid Tritylodon and possible saurischian dinosaurs. The clearly associated, articulated ornithischian material includes a series of posterior dorsals, sacrals and anterior caudals, pelves and partial hindlimbs ( Fig. 5 View Figure 5 ). The length of the ilia and femora indicate that, although this is a relatively small individual, this material is of a slightly larger animal than the syntypes of Lesothosaurus (BMNH RUB17, RUB23). A number of features of this material suggest immaturity: although usually closely articulated the neurocentral sutures are not fused, the sacral and caudal ribs are separate and the sacral vertebrae are not fused (but as noted above, lack of vertebral fusion may be a persistent feature of basal ornithischians).
The ischium of BP/1/4885 clearly has a well-defined, tab-like obturator process ( Fig. 5 View Figure 5 ), which is comparable to that of SAM-PK-K1105 and BMNH R11000; this process is not seen in Lesothosaurus ( Figs 3A–B View Figure 3 , 4 View Figure 4 ; see above). Although Knoll (2002a, b) considered BP/1/ 4885 to be an individual of Lesothosaurus , no characters (other than perhaps the relatively small size) support this referral. With the exception of the obturator process, the rest of the preserved material is morphologically indistinguishable from both Lesothosaurus and Stormbergia . Thus, BP/1/4885 cannot be used to infer the presence of an obturator process in Lesothosaurus diagnosticus ; the only specimen that can legitimately be used for that purpose is the syntype BMNH RUB17, and this specimen (as noted above) demonstrates that an ornithopod-like obturator process is clearly absent. The ornithischian material in BP/1/ 4885 is here considered to represent a juvenile individual of Stormbergia based upon the presence of the tab-shaped obturator process.
BP/1/4885 further supports the hypothesis that the Lesothosaurus diagnosticus syntypes (BMNH RUB17, RUB23) are not juveniles of Stormbergia : a wellformed obturator process is seen in an individual that is only 20% larger than BMNH RUB17. Evidence from a growth series of Hypsilophodon (BMNH R195, R193, R5830) suggests that the obturator is fully formed in relatively young individuals and does not change significantly during ontogeny.
The features that distinguish Stormbergia from Lesothosaurus are not autapomorphies. A tab-shaped obturator process is seen in some basal ornithischians (e.g. Agilisaurus, Peng, 1997; ‘ Y. ’ multidens, He & Cai, 1984) and ornithopods (e.g. Hypsilophodon, Galton, 1974a ), while the loss of an elongate ischial symphysis characterizes nearly all ornithischians. However, a combination of plesiomorphic and derived features allow Stormbergia to be distinguished from all other basal ornithischians, as in Lesothosaurus .
Several derived postcranial features are present which distinguish Stormbergia from Pisanosaurus . These include the presence of an opisthopubic pubis, development of a prepubic process, and the well-developed lateral flange of the distal tibia, all of which are absent in Pisanosaurus ( Sereno, 1991) . The presence of an untwisted ischial shaft, with a well-developed, tab-shaped obturator process, distinguishes Stormbergia from an undescribed basal ornithischian from the Late Triassic Lower Elliot Formation of South Africa (SAM-PK-K8025). Although cranial material of Stormbergia is unknown, postcranial anatomy distinguishes Stormbergia from the heterodontosaurs Heterodontosaurus and Abrictosaurus . Both these taxa have the derived conditions of a horizontal (and greatly reduced) brevis shelf, a fully open acetabulum and the absence of a supraacetabular flange. Additionally, a tab-shaped obturator process is absent from the ischium of Heterodontosaurus . Finally, Stormbergia can be distinguished from Agilisaurus and ‘ Y. ’ multidens by the retention of an elongate pubic peduncle of the ilium and a short prepubic process of the pubis, and additionally from ‘ Y. ’ multidens by the retention of a supraacetabular flange and ventromedially angling brevis shelf on the ilium.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Fabrosaurus australis
| Butler, Richard J. 2005 |
Scutellosaurus lawleri
| Colbert 1981 |
LESOTHOSAURUS DIAGNOSTICUS
| GALTON 1978 |
Pisanosaurus mertii
| Casamiquela 1967 |