
Iberolacerta galani, Arribas, Oscar, Carranza, Salvador & Odierna, Gaetano, 2006
|
publication ID |
https://doi.org/10.5281/zenodo.172862 |
|
DOI |
https://doi.org/10.5281/zenodo.6255230 |
|
persistent identifier |
https://treatment.plazi.org/id/03FE4B1B-FF9F-B961-1A17-FD8468CBFC5F |
|
treatment provided by |
Plazi |
|
scientific name |
Iberolacerta galani |
| status |
sp. nov. |
Iberolacerta galani sp. nov.
[ Fig. 8 View FIGURE 8 A–H; Fig 9 View FIGURE 9 A–D]
Synonymy
First nomenclatorial combinations, which include specimens from Iberolacerta galani nov. (localities originally referred to are in parenthesis).
Lacerta monticola cantabrica (partim); Elvira & Vigal, (1982), Doñana, Acta Vertebrata, 9: 100, fig 3 (from Truchillas, León and Laguna Vega de Porto, Zamora).
" Lacerta " monticola (partim); Arribas, (1996), Herpetozoa, 9 (1/2): 32 (from Truchillas, León).
Lacerta (Iberolacerta) monticola cantabrica (partim); Carranza et al. (2004), Systematics & Biodiversity 2(1): 61 (from Laguna de Los Peces, Zamora).
Holotype
IPE 4000 Adult male ( Fig.7 View FIGURE 7 and 8 View FIGURE 8 A–B). Red plastic label attached to left forelimb with engraved number IPE4000. Also, white cardboard label attached to the left forelimb. Anverse (handwritten): "Laguna de los Peces (Zamora). Sª Segundera. 1700 m. 25804. 2 (encircled). Oscar Arribas". Reverse (handwritten): " Iberolacerta galani sp. nov. ". A red plastic label attached to left hindlimb (in white letters and relief) " HOLOTYPUS ". In the I.P.E. collection (Instituto Pirenaico de Ecologia, Jaca, Spain, belonging to the C.S.I.C.).
Teleno Trevinca Cabrera Sanabria
(Morredero & Portillinos) (Fonte da Cova) (Lago de Truchillas) (Laguna de los
Peces) 19.19–21.76 15.97–20.52 15.87–24.9 15.4–23.8
HLL 29.1±1.58 26.87±3.18 32.34±1.30 29.50±0.90 Teleno Trevinca Sanabria
(Morredero & Portillinos) (Fonte da Cova) (Laguna de los Peces)
n=9 n=3 n=14
SVL 58.97±2.5 69.04±3.73 66.51±2.9
48.25–72.18 62–74.71 52.75–84.42
FLL 18.38±0.63 20.01±0.83 19.96±0.62
15.18–20.42 18.42–21.26 16.25–24.05
HLL 25.86±0.70 28.79±1.13 28.04±0.77
22.75–29.54 26.63–30.48 24.28–32.98
PL 12.75±0.37 14.16±0.58 13.88±0.41
11.27–14.83 13–14.9 11.8–16.65
PW 6.29±0.14 7.1±0.32 6.73±0.18
5.6–6.86 6.5–7.6 5.7–7.9
PaL 4.19±0.13 4.83±0.27 4.63±0.17
3.7–4.93 4.3–5.2 3.7–5.56
DM 1.85±0.09 1.63±0.07 1.75±0.14
1.57–2.44 1.49–1.73 0.86–2.58
DT 1.9±0.12 1.98±0.15 1.94±0.04
1.41–2.5 1.68–2.22 1.36–2.63
AW 3.45±0.24 3.97±0.08 3.84±0.23
2.15–4.86 3.88–4.14 2.58–5.28
AL 2±0.11 2.31±0.15 2.27±0.12
1.52–2.53 2.02–2.52 1.79–3.29
GrS r 8.66±0.70 9.66±1.85 10.28±0.45
6–11 6–12 7–13
GrS l 8.77±0.40 9.66±1.45 10±0.46
7–11 7–12 5–12
GUL 24.66±0.60 23.66±0.88 23.42±0.67
23–29 22–25 16–27
COLL 11.77±0.43 11.33±0.66 10.85±0.4
10–14 10–12 9–14
DORS 52.88±0.88 53±1.15 53.28±1.11
48–56 51–55 48–61
to be continued Paratypes
Eight males and thirteen females from Laguna de los Peces, Sierra Segundera, ca. 1700 m, same data as Holotype (IPE 4001–4005 and OA 04082501, 3–18). One male from Lago de Truchillas, Sierra de la Cabrera Baja (León province), 12/04/1990 (nº OA90041201). Eight males from Lago de Truchillas, Sª de la Cabrera Baja, 1390–1750 m (León province), 6/04/1997 (nº OA97040601–08). Six males and five females from Puerto del Morredero (ski resort), Sierra del Teleno, 1762 m (León province), 14/08/2005 (nº OA 05081401 –09, OA 05081412, OA 05081416). Two males and four females from Puerto Los Portillinos, Sierra del Teleno, 1820–1957 m (León province), 14/08/2005 (nº OA 05081401 –06). Two males and three females from “Refugio da Fonte da Cova” (abandoned ski resort), Peña Trevinca, 1860 m (Orense province), 15/08/2005 (nº OA 05081501 –05). All with plastic red labels attached to their left hindlimbs (with white letters in relief reading " PARATYPUS ").
Diagnosis
A large Iberolacerta , especially characterized by the following combination of characters: large SVL (females up to 84.42 mm, the largest Iberolacerta known to date) with relatively small fore and hindlimbs. High number of blue occelli on the shoulders (UVreflective, as are the blue spots on the outermost ventral ranges, see Figs. 8 View FIGURE 8 and 9 View FIGURE 9 ) and contact between supranasal and first loreal contact relatively frequent (full contact in almost a quarter of all specimens analyzed, and near contact in many others; Fig. 7 View FIGURE 7 ). Frequency of rostralinternasal contact relatively low in males (33 %) but relatively high in females (58 %), higher number of Collaria, Dorsalia and Ventralia and less Circumanalia in comparison with other Iberolacerta . Postocular and parietal plates separated. Azygos scale between the prefrontals rare (13 % of specimens). Osteologically, it is characterized by a fairly straight squamosal bone, only incurved in its posterior part ( Fig. 6 View FIGURE 6 ), Karyotype with 36 uniarmed machrochromosomes gradually decreasing in length and NORs in the telomeric position of a large chromosome pair (possibly the fifth chromosome pair; LType) ( Figs 4 View FIGURE 4 and 5 View FIGURE 5 ). Sex chromosomes differentiated and heteromorphic. Chromosome Z presents a peritelomeric Chromomycin A3 heterochromatic band, which is unique among Iberolacerta and is as large as the autosome pair 6. Chromosome W is heterochromatinized and as large as autosome pair 10 or 11. Partial mitochondrial DNA sequences for the cytochrome b and 12S rRNA genes sequenced for this study are distinct from all the remaining representatives of Iberolacerta known to date ( Fig. 3 View FIGURE 3 ).
Description of holotype
Biometry: Adult male with snoutvent length of 69.05 mm. Tail 29.16 mm (autotomized). Forelimb length 23.84 mm. Hindlimb length 33.06 mm. Pileus length 16.8 mm. Pileus width 7.5 mm. Parietal legth 6.16 mm. Masseteric widest diameter 2.54 mm. Tympanic widest diameter 1.87 mm. Anal plate width 4.39 mm. Anal length 2.56 mm.
FLL/SVL (relative forelimb length): 0.345. HLL/SVL (relative hindlimb length): 0.478. PL/PW (pileus shape): 2.252. DM/PaL (relative masseteric plate size): 0.412. DT/PaL (relative tympanic size): 0.303. AL/AW (anal plate surface): 0.583. AS /SVL (relative anal plate size with respect to total length): 4.854.
Scalation
Number of supraciliary granules: 10 (right) and 9 (left). Supralabials: 4 (both sides). Sublabials: 7 (both sides). Submaxillars: 6 (both sides). Gularia: 22. Collaria: 9. Dorsalia: 55. Ventralia: 27. Femoral pores 17, on both sides. Lamellae: 25. Circumanal Plates: 6. Scales on a ring annulus from tailbasis: 27. Rostral in full contact with internasal. Supranasal in contact with first loreal. One postnasal. First postocular separated from parietal plate. An azygos (supernumerary) scale between prefrontals. Occipital and interparietal plates separated by a prolongation of the right parietal, and a supernumerary small scale corresponding to this same anomalous prolongation on the left side. An illustration of a lateral view of the head of the holotype is shown in Fig. 7 View FIGURE 7 .
Coloration (in life, outside the breeding period)( Fig 8 View FIGURE 8 A–B). Pileus with tiny, vermiculated irregular spotting. Small spots on supralabials, subocular and sublabials. Small spots on the sides of the gular area. Dorsal tract with yellowishgrey (4B2) [2 Y 8.1/ 1.3] to greyishyellow (4B3) [3.5 Y 8.0/2.6] base color in life (same color as in alcohol). Two juxtaposed rows of black spots fused together forming transverse irregular spots that cover approximately half of the dorsal tract width. These black spots get smaller and fainter along the tract towards the tail. Temporal and infratemporal bands fused and reticulated, leaving small whitish (or blue, see below) spots, more developed in the area within these two primitive bands, on the lower part of the flanks. This temporal band starts at the eyes and runs along the sides, where it narrows and appears faintly on the sides of the tail. Four (right side)/five (left side) blue (21A7) [6PB 5.0/12.4] occelli on the shoulders. Small blue dots on the outermost plates of the venter. Venter pastel green (29A4) [6gy 8.7/3.1] on the belly and greenish white towards the limits of the gular area (29A2) [4GY 9.0/1.0] in life. In alcohol, from turquoise white (24A2) [5B 8.7/1.0] on the central plate rows of the venter to light turquoise (24A4) [6.5B 7.7/3.1] towards the outermost ones. The four outermost ventral plate ranges with black spotting (the two outermost well developed, sinuose and placed on the foreborder of the plate, covering near half of its surface; the two inner plate ranges only thin and on the foremost border). Two minute spots on the posterior free border of anal (preanal) plate. Blue occelli on the shoulder UVreflective, as are the blue spots on the outermost ventral ranges.
Variability
Biometric and scalation values for the whole species, and comparison with the other Iberolacerta species from the “monticolagroup” are shown in Tables 5 View TABLE 5 (males) and 6 (females). Intraspecific variability of I. galani sp. nov. by samples is presented in Tables 8 View TABLE 8 (males) and 9 (females) (only specimens with SVL> 45 mm). The small size of samples precludes any statistical comparisons between them. Pictures of I. galani nov. (fig 8A–H & 9A–D) and some I. monticola are shown for comparison ( Fig. 10 View FIGURE 10 , A–H).
Concerning certain singular morphological characters, of nineteen male specimens, fifteen exhibit the common disposition of prefrontals in full contact, whereas two specimens have an azygos scale between them, and another two show separation between these scales, permitting a suture between internasal and frontal plates. Out of twentysix females, twentythree exhibit the common disposition of prefrontals in full contact, whereas three have an azygos scale between them. Nine specimens from a total of fortyfive (including both sexes) have two masseteric plates instead of one.
Coloration males (breeding period, from Sierra de la Cabrera). In life, dorsal tract from green (28B7 to 28B6) [7.5GY 7.0/7.8 to 7.5GY 7.3/5.7] to greenish grey (28B2) [5GY 8.0/ 0.8], and to greyish green (28C4) [6GY 6.8/2.5] in the pelvic area and forelimbs. Shoulder occelli vivid blue (22A8) [5PB 4.5/14.2], very numerous, in most cases from two to seven but occasionally up to eleven ( Fig 9 View FIGURE 9 D). The irregular vertebral band (more barely aligned in two juxtaposed rows) is very developed and finely disgregated, connected to the temporal (=costal) bands giving the specimens a highly reticulated overall appearance. This band connects rapidly and progressively with the costal ones as the specimens grow. Ultraviolet reflective occelli are present in I. monticola , I. martinezricai and I. galani nov. In this latter species ( Fig 8 View FIGURE 8 E–F), however, blue (and UV reflecting) occelli are more numerous (and are absent in Podarcis bocagei , syntopic in some localities). The number and size of UV reflecting occelli increases with age and, presumably, status.
Females (breeding period). Unknown.
Males (outside the breeding period, from Sierra Segundera, Trevinca and Teleno) ( Figs 8 View FIGURE 8 A, B and E). As in the holotype, yellowishgrey (4B2) [2 Y 8.1/1.3] to greyishyellow (4B3) [3.5 Y 8.0/2.6] dorsal tract base color, only slightly darker in specimens close to ecdysis. Specimens that are not yet fully grown (especially those living on slates in Teleno and Trevinca) have a brownish dorsum. Vertebral band with small or medium irregular dots, even elongated and vermiculated, which are barely aligned in two juxtaposed (but distinguishable) rows. These dots or irregular blotches tend to coalesce with age (as in the holotype). Temporal (=costal) bands reticulated, with scalloped upper edge and also fused with the "lateral inferior line", leaving a row of clearer dots among the primitive situation of both, which are a prolongation of the blue occelli of the shoulders. Venter pastel green (29A4) [6gy 8.7/3.1] on the belly and greenish white towards the limits of the gular area (29A2) [4GY 9.0/1.0] in life. The occelli on the shoulders are blue (21A7)[6PB 5.0/12.4]. Usually only the two outermost rows of ventral plates present dark marks. These appear more centrally on the plate in younger males but are bigger and connected to the foreborder in older males. There are a few black dots on the submaxillar plates and especially in the gular sides.
Females (outside the breeding period, from Sierra Segundera, Trevinca and Teleno): ( Figs. 8 View FIGURE 8 C, D, G and H). Dorsum from pale green (29A3) [6GY 8.9/2.1] to greyish green (29B3 and 29B4) [5GY 8.0/1.8 and 5.5GY 7.9/2.8) in fullygrown animals. Pattern is much less spotted than on males and the degree of development is not correlated with age (older females may or may not exhibit moderately developed spotting). Usually a dark, irregular spotting or vermiculated in the center of the dorsum, which can either be spread widely across the dorsal tract, or be clearly aligned in two rows. Temporal bands not reticulated but uniform, with marginal areas (especially the upper border) darker (nearly black) and the inner area lighter (brown). Upper edge of this temporal (=costal) band also scalloped, enclosing lighter dots, more visible in younger specimens but also in adults. The rows of lighter dots on the lower part of the sides are less marked, as is the inferior lateral line, which is faint and rarely appears in the form of dots. Venter yellowish green (30A8 and 30A7) [3.5GY 8.5/11.4 and 3.5 8.7/8.6]. Blue occelli as in males. In contrast to the males, black spotting usually appears in the center (or on the rear parts) of the plates of the outermost ventral scale rows, and is often faint. Less frequently, very small dots appear on the second inner row of ventral plates.
A melanic female (the first published case of melanism in the I. monticola group) is depicted in Fig 9 View FIGURE 9 C.
Hatchlings (from Sierra Segundera and Teleno): ( Figs. 9 View FIGURE 9 –A–B). Dorsum with greyishyellow (2B3 and 2B4) [8 Y 8.0/2.5 to 9 Y 8.0/4.1] background. Dorsal tract finely spotted with irregular blotches on the vertebral area, which can be very faint and barely distinguishable. Temporal bands slightly reticulated in male hatchlings and more uniform in females (sexual dimorphism from birth). Venter without color pigmentation, with the first (outer) row of ventrals densely spotted, but the inner second and third pairs of rows less marked. Tail is blue (22B7 and 23B7) [5PB 4.9/11.0 and 1PB 5.1/9.2].
Additional material. Additional unnumbered specimens were cleared and stained for the osteological (two from Sanabria) and karyological work (three: two from Sanabria and one from Teleno). Additionally, nine live hatchlings were examined (six from Sanabria and three from Teleno). Almost twenty other adult specimens were examined and released in Sanabria, four adults from Teleno and another subadult from Trevinca.
COMPARATIVE NOTES: (taxa with significant differences to I. galani nov. in parenthesis; abbreviations, cyr: I. cyreni ; mart: I. martinezricai ; mont: I. monticola ).
Morphology
MALES of I. galani have fewer left supraciliary granules (cyr), greater collaria (cyr), greater ventralia (cyr), small left femoralia (mart), less circumanalia (cyr), low frequency of rostralinternasal contact (cyr, mont), greater supranasalloreal contact (all other), higher number of blue occelli (all other), greater ventral punctuation (cyr), shorter forelimb length (cyr, mont), shorter parietal length (cyr), greater masseteric plate (cyr, mart), greater tympanic (cyr) and small anal plate (cyr).
FEMALES of I. galani nov. have less fewer supracilliary granules (cyr), greater collaria (all other), greater dorsalia (cyr, mont), greater ventralia (all other), less circumanalia (cyr, mont), low frequency of rostralinternasal contact (cyr) supranasalloreal contact (all other) higher number of blue occelli (all other), ventral punctuation (cyr), shorter forelimb length (cyr), shorter hindlimb length (cyr, mont), shorter pileus (cyr), greater masseteric plate (mart), greater tympanic size (all other) and smaller anal plate (cyr).
The presence of an azygos scale between prefrontals is rare in I. galani nov. (13 %), whereas it is very common in the immediate Galician populations of I. monticola (present in more than 50% of the examined specimens from most Galician populations and in all of the examined specimens from the Cabeza de Manzaneda (Orense), the nearest known population to the general area of I. galani nov., less than 40 km in a straight line; see Fig. 1 View FIGURE 1 ).
Osteologicaly, I. galani nov. is very similar to the other species in the “Iberian group” (see Fig. 5 View FIGURE 5 ). However, it differs in the shape of the squamosal bone, which is clearly curved in all the other Iberolacerta (including the Pyrenean and Alpine species) but fairly straight in I. galani nov. (see Fig. 6 View FIGURE 6 ). Moreover, in the “Iberian group”, I. cyreni usually has longer nasal bones and nine premaxillary teeth (seven in the I. galani nov. studied); I. monticola has a spatuliform or elongated and parallelsided processus nasalis (only rarely arrowshaped, more frequent in Galicia and Estrela, also present in I. galani nov.); I. martinezricai has nine premaxillary teeth, an elongated (parallelsided, not arrowshaped as in I. galani nov.) processus nasalis, and frequently seven short presacral ribs and a thin triangular medial process in the squamosal, at least in some specimens.
The karyotype of I. galani nov. is distinguished from the karyotypes of I. martinezricai and I. cyreni by the position and location of the NORs (telomeric and on a long chromosome pair in I. galani nov. and interstitial and on a mediumlarge sized pair in I. martinezricai and I. cyreni ). It also differs from the karyotype of I. monticola by the presence of differentiated sex chromosomes, Z and W, both heteromorphic and heterochromatic (homomorphic and euchromatic in I. monticola ) (see Fig 5 View FIGURE 5 ). It differs from I. horvathi (which also has partially heterochromatic W sex chromosome and LType NORs) by the fact that in I. galani nov., W is fully (not partially) heterochromatic and mainly by the presence of a Z, unique among Iberolacerta , in bearing a peritelomeric CMA3 positive heterochromatin.
Derivatio nominis
The specific epithet " galani " is a genitive (possessive) Latin name (g. masculine). This patronym (eponym) honours Dr. Pedro Galán Regalado (Cee, 1955), from A Coruña ( Spain), for his lifelong dedication to the study of NW Iberian (especially Galician) herpetofauna and Natural History in general. This is also in recognition of the discovery and longterm study of most of the lowland and sealevel populations of Iberolacerta monticola from Galicia.
Distribution
Iberolacerta galani nov. is endemic to the Montes de León (northwestern Iberian Peninsula, see Fig. 1 View FIGURE 1 ): Sierra Segundera, Sierra de la Cabrera, Sierra del Eje or Peña Trevinca and Sierra del Teleno between the provinces of León, Zamora and Orense, where it can be found from 1000 to 2000 m, but probably also at higher altitudes. The first two mountain ranges encircle the highland plateau of Sanabria, where as a result of Quaternary glacial activity many lakes were formed. Nearby, I. galani nov. is present in the Sierra del Eje or Peña Trevinca, on the border between the provinces of Orense and León, where it has been found from 1700 to 2000 m. It is also present to the north of the Eria and Cabrera rivers, in the neighboring Sierra del Teleno in the province of León, where it has been recorded from 1340 to almost 2000 m, but where it probably reaches the highest peaks of these ranges (Vizcodillo 2122 m, Trevinca 2096 m, and Cerro del Teleno 2185 m) (see Figs. 9 View FIGURE 9 E, F, G and H).
The mountain area inhabited has the form of a “V” without vertex, open to the west, formed by the anticlinals of PiornalTeleno (to the North) and Ollo de Sapo (to the South) and separated in the middle by the synclinal of Truchas. The overall distribution range of I. galani nov. borders several lowland areas unsuitable for mountain lizards like Iberolacerta . More specifically, its distribution borders with El Bierzo (Sil valley) to the north; with La Maragateria (Duero Valley) to the north east and east; with the Tera valley (draining into the Duero) to the south; and with the Bibei river valley (draining into the Sil) to the west. These lowland areas have either a moderate Atlantic (to the north) or Mediterranean (the remainder) climate ( NietoFeliner 1985). Further west, on the other side of the dry Mediterranean Bibei river valley, I. galani nov. is replaced by I. monticola , which inhabits the Serra da Queixa (summit in Cabeza Grande de Manzaneda— 1778 m) and the Serra do Invernadeiro (both in Orense province). These I. monticola populations lie less than 40 km away from the nearest I. galani nov. populations from Peña Trevinca, also in the province of Orense (see Fig. 1 View FIGURE 1 ).
Iberolacerta monticola is also found very close to the northern part of the distribution range of I. galani nov., on the other side of the Sil river valley in the Sierra de Ancares, the Sierra de Caurel, etc. and further to the north east, in the mountainous regions leading to the main axis of the Cordillera Cantabrica (see Fig. 1 View FIGURE 1 ). The highest pass between the Montes de León (inhabited by I. galani nov.) and the spurs of the Cordillera Cantabrica (inhabited by I. monticola ) is the Puerto del Manzanal ( 1230 m), where Iberolacerta has never been recorded.
Habitat
Within its distribution area, Iberolacerta galani nov. inhabits supraforestal habitats, which are characterized by a highmountain climate and are included in the Oromediterranean and Crioromediterranean bioclimatic stages ( Rivas Martinez 1987) (see Figs 9 View FIGURE 9 E, F, G and H).
The rocky substrates inhabited are fairly diverse: in Sanabria and the Sierra de la Cabrera it lives upon a small number of fissured igneous rocks (“Ollo de Sapo” and other sincynematic granitoid rocks of uncertain age) ( Fig 9 View FIGURE 9 G); in Teleno ( Fig 9 View FIGURE 9 E and F) and Trevinca ( Fig 9 View FIGURE 9 H) it lives among slabs of black slate from the Middle Ordovician age, highly prized for their use in construction, which could represent a threat to its conservation in some areas; in the highest parts of these sierras (Teleno and Cabrera) it lives among the hard Armorican quartzites that constitute the peaks.
The lower areas currently inhabited by I. galani nov. may well have been colonized by the species in historical times, when the forest was cleared for pastures. The species can also inhabit areas of azonal vegetation below its natural limit, in particularly favourable conditions in terms of temperature and moisture such as river canyons. These lower areas are included in the Supramediterranean and altimontane Estrellian, OrensanoSanabrian and Galaicoportuguese Silicicolous Series of Betula celtiberica or birch ( Saxifrago spathularidiBetuleto celtibericae sigmetum) and are constituted by the upper limits of the forests, usually destroyed as a result of fires. The regressive stages inhabited by I. galani are broom heathlands of Cytiso striati Genistetum poligaliphyllae, which are themselves replaced by heathlands of Genistello tridentatae Ericetum aragonensis ( NavarroAndrés & ValleGutierrez 1987; Rivas Martinez 1987).
The main part of the I. galani nov. distribution range falls between the Oromediterranean OrensanoSanabrian, silicicolous series of the dwarf juniper ( Genisto sanabrensis Junipereto nanae S.) which are areas with silicicolous, psicroxerophilous and chionophilous oromediterranean shrubs, endemic to the high OrensanoSanabrian Sierras and belonging to the Genisto sanabrensis Juniperetum nanae, which is its climax series. Covers from 1600m to 2000m. A shrub endemic to the area is Genista sanabrensis ValdesBermejo, Castroviejo & Casaseca, 1977 . Trees are absent from these zones, and the dense scrub has Juniperus nana , Genista sanabrensis, Cytisus oromediterraneus (= C. purgans ) and Deschampsia iberica . The more degraded scrub has Calluna vulgaris, Cytisus oromediterraneus, Genista sanabrensis and Erica aragonensis . Grasslands contain Nardus stricta, Agrostis capillaris and Phalacrocarpum oppossitifolium ( NavarroAndrés & ValleGutierrez 1987; Rivas Martinez 1987).
The upper parts of its altitudinal range (above 2000 m) belong to the Crioromediterranean OrensanoSanabrian silicicolous series of Festuca indigesta ( Teesdaliopsio confertae Festuceto indigestae S.), which appears only on those summits above 2000 m, where it forms a mosaic with dwarf shrubs of Genisto sanabrensis Junipereto nanae S. ( NavarroAndrés & ValleGutierrez 1987; Rivas Martinez 1987). An exhaustive account of the flora from this area can be found in NietoFeliner (1985).
Interestingly, these areas have undergone dramatic changes in their vegetation during the Holocene. Conifers ( Pinus sylvestris and Pinus uncinata ), today absent, were present in the Cordillera Cantabrica, Galicia and the Montes de León during the Pleistocene and well into the Holocene, but disappeared very recently (probably during the Subatlantic period, less than 2500 ybp) as a result of oceanic conditions and human influence (there are no conifers in the Cantabric subalpine belt).
Thanks to one of the best pollinic sequences of the Iberian Peninsula (Laguna de las Sanguijuelas, Sanabria, 1000 m, with a time span of nearly 14000 years), the vegetation history of Sanabria is extremely well known and indicates that conifers ( Pinus sp.) were well represented until very recently and only retreated during Subatlantic, when they were substituted by Quercus at low altitude and Betula at high altitude ( Blanco et al. 1996; Arribas 2004).
Endemic zoological species from the Montes de León include mainly coleopters with dispersive capabilities very similar to those of mountain lizards: Nebria belloti Franz, 1954 (Trevinca, Segundera and Teleno) , Nebria leonensis Assmann, Wrase & Zaballos, 2000 (Cabrera baja and Teleno), Leistus valcarceli Wrase, RuizTapiador & Zaballos, 1998 (Cabrera Baja and Teleno) , Zabrus mateui Novoa, 1980 (Peña Trevinca, Sierra Cabrera Baja, Sierra Segundera and Montes Aquilanos) and Anchomenidius feldmanni Wrase & Assmann, 2001 (Teleno) ( Alonso et al. 1987; Wrase et al. 1998; Assmann et al. 2000 and our own data). Most of these new species are recent discoveries and, like the lizards of the I. monticola group, they belong to the biogeographical “Lusitanian” species groups that are distributed throughout the western Iberian Peninsula, and which spread further into the center and the east along the main mountain ranges, with a high incidence of endemics ( Arribas & Carceller 1995; Arribas 2004).
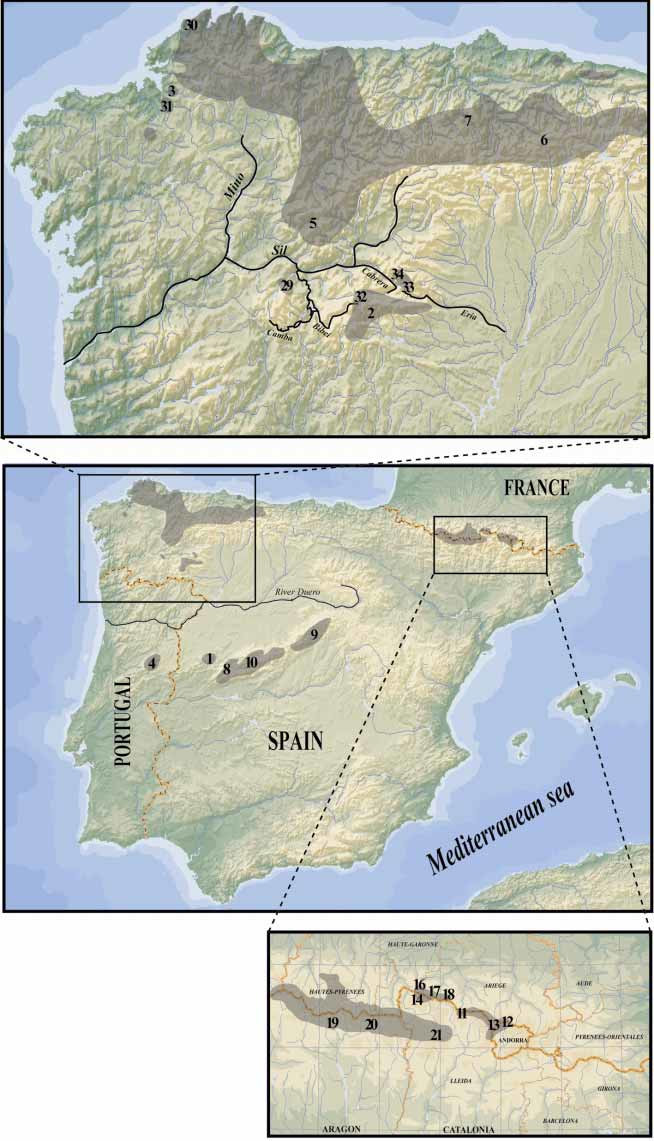
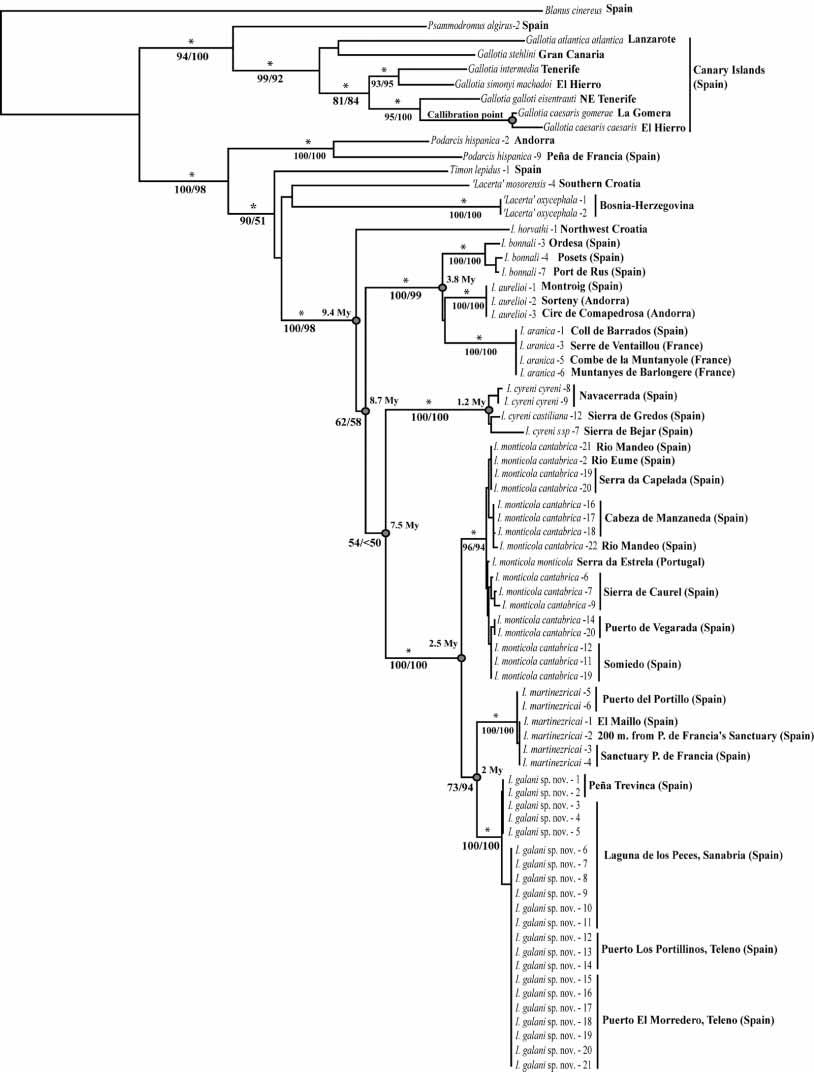



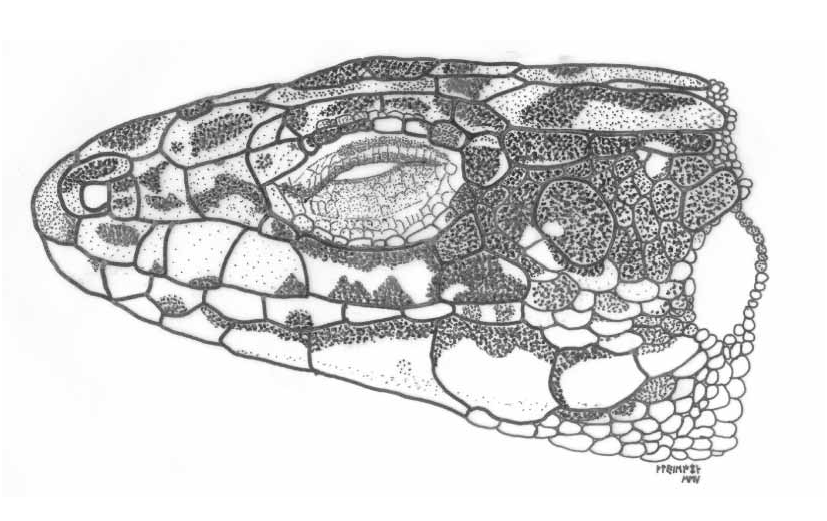



TABLE 8. Morphometric, scalation and biometric indexes of Iberolacerta galani nov. males from different localities. See text for abbreviations of characters and indexes used in the morphometric analysis.
| n=3 | n=2 | n=9 | n=10 | |
|---|---|---|---|---|
| SVL | 58.47±4.15 | 53.65±7.82 | 63.28±2.22 | 60.66±1.84 |
| FLL | 53.72–66.75 20.30±0.76 | 45.83–61.48 18.24±2.27 | 47.11–69.1 22.33±0.91 | 51.56–69.05 19.92±0.88 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |