
Lebbeus thermophilus, Komai, Tomoyuki, Tsuchida, Shinji & Segonzac, Michel, 2012
|
publication ID |
https://doi.org/10.5281/zenodo.280458 |
|
DOI |
https://doi.org/10.5281/zenodo.5490704 |
|
persistent identifier |
https://treatment.plazi.org/id/03FB87A2-7D58-DC2D-F483-FDC7FAEFFC6A |
|
treatment provided by |
Plazi |
|
scientific name |
Lebbeus thermophilus |
| status |
sp. nov. |
Lebbeus thermophilus sp. nov.
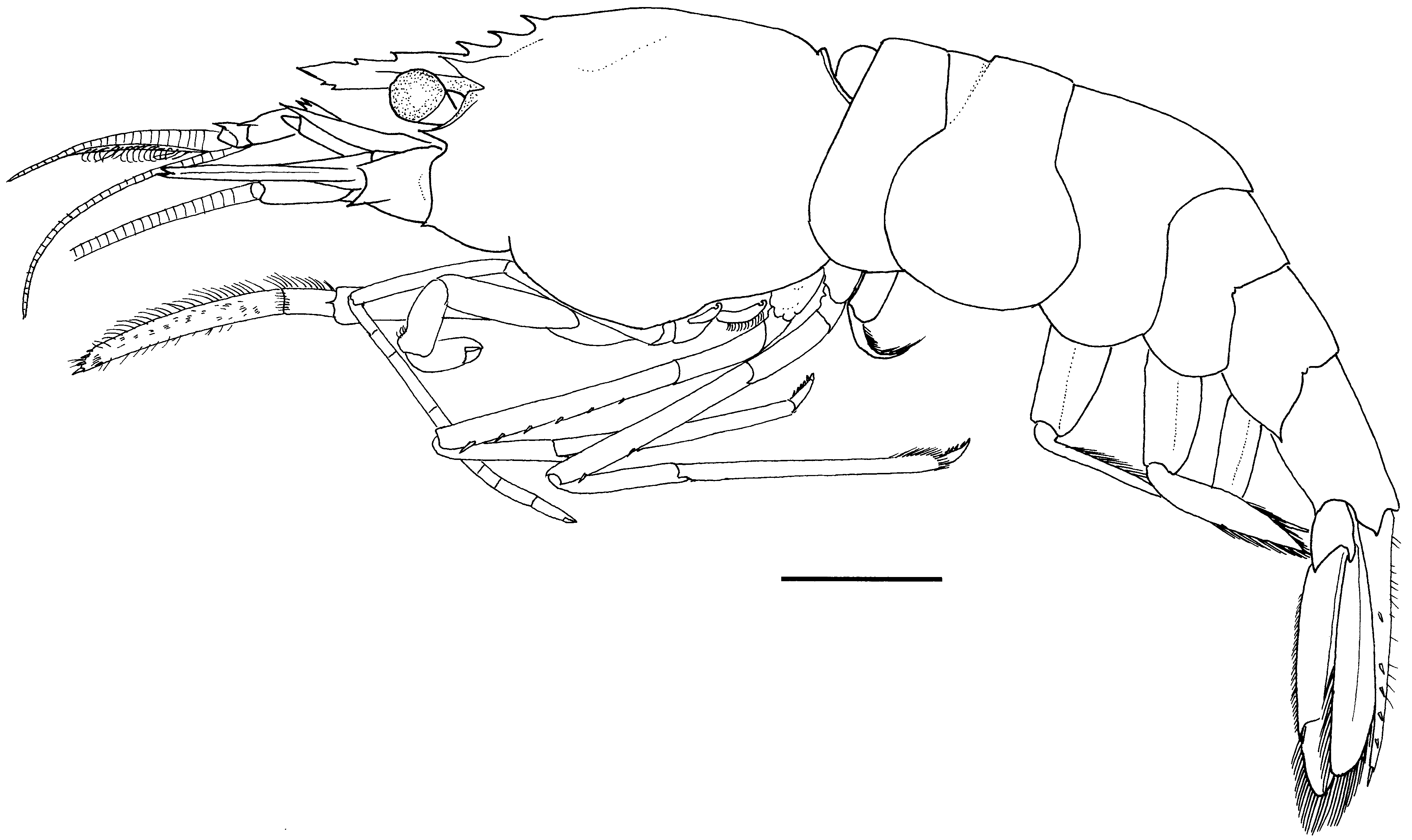
( Figs 14–17 View FIGURE 14 View FIGURE 15 View FIGURE 16 View FIGURE 17 )
Lebbeus sp. aff. washingtonianus: Komai & Segonzac 2004: 1179 . Lebbeus sp. A: Martin & Haney 2005: 449 View Cited Treatment , table 1.
Material examined. Holotype: RV Natsushima /DS S hinkai 2000, NT99-15 cruise, dive #1150, PACMANUS , Manus Basin, Bismarck Sea, 03°43.267’S, 151°40.480’E, 1662 m, 7 November 1999, female (cl 11.3 mm), NSMT-Cr 21984 (ex-JAMSTEC 019523–019525).
Paratypes: Manus Basin, Drill Ship Hunter /ROV TST212 Canyon Offshore, dive #0011, Suzette Vent Field, Binns Mound, 03°58.1168’N, 152°93.65’E, 1512 m, June 2005, 1 ovigerous female (cl 16.1 mm), MNHN-IU- 2011-5720; 2007 Wave Mercury cruise, dive #36, site South Su, 03°50.97’S, 152°10.53’E, 1323 m, 16 April 2007, 4 females (cl 10.5–19.0 mm), 1 male (cl 9.8 mm), CBM-ZC 10661; dive #38, same site, 16 April 2007, 4 females (cl 9.0–18.0 mm), CBM-ZC 10662.
Lau Basin, BIOLAU, BL 0 2, site Hine Hina, 22°32'S, 176°43'W, 1842 m, 14 May 1989, baited trap, 1 female (cl 19.2 mm), MNHN-IU-2011-5140; BL 0 3, same site, 15 May 1989, baited trap, 16 females (cl 7.6–18.4 mm), 2 ovigerous females (cl 15.1, 15.8 mm), 2 males (cl 9.0, 9.5 mm), MNHN-IU-2011-5719; BL 0 6, 1 juvenile (cl 5.0 mm), MNHN-IU-2011-5145; BL 0 8, site Vailili, 23°13'S, 176°38'W, 1740 m, 18 May 1989, slurp gun, 4 females (cl 8.8–15.8 mm), 1 male (cl 9.5 mm), 1 juvenile (cl 5.9 mm), MNHN-IU-2011-5139; same data, 2 females (cl 13.0, 17.2 mm), MNHN-IU-2011-5141; BL 10, same site, 22 May 1989, slurp gun, 2 females (cl 13.1, 14.3 mm), MNHN-IU-2011-5142; BL 11, same site, 23 May 1989, slurp gun, 1 female (cl 7.3 mm), MNHN-IU-2011-5144; BL 12, same site, 24 May 1989, baited trap, 1 female (cl 10.7 mm), MNHN-IU-2011-5143.
Description. Females. Body ( Fig. 14 View FIGURE 14 ) moderately robust for genus; integument moderately firm, surface glabrous.
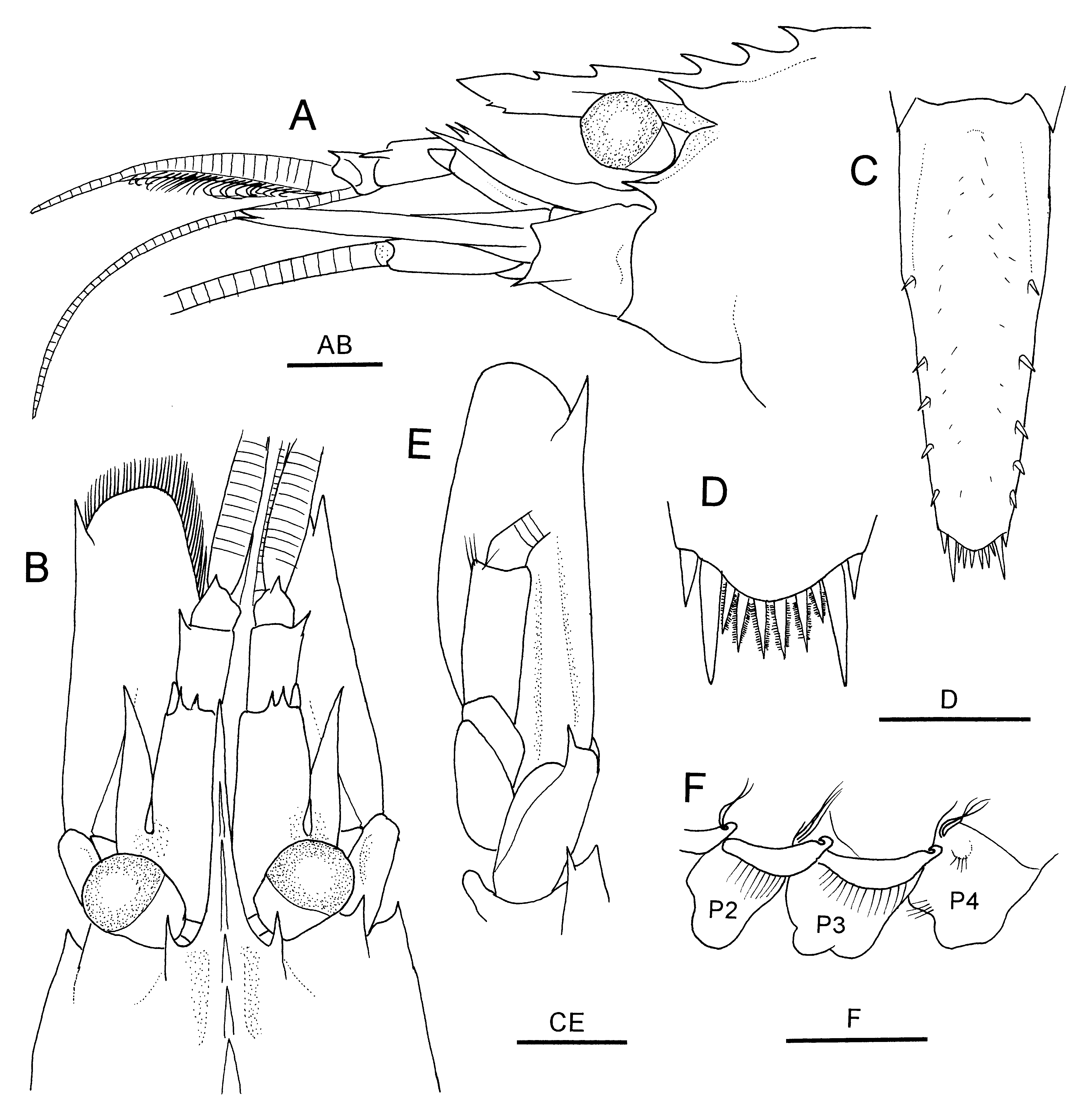
Rostrum ( Figs. 14 View FIGURE 14 A, 15A, B) straight, directed forward, reaching or slightly falling short of distal margin of first segment of antennular peduncle, 0.4–0.5 times carapace length; dorsal rostral series consisting of 5 teeth, including 2 small teeth on rostrum proper and 2 or 3 small to moderately small postrostral teeth, ventral margin armed with 2 tiny subterminal teeth, ventral lamina slightly developed. Carapace ( Figs. 14 View FIGURE 14 , 15 View FIGURE 15 A, B, 17D) with low postrostral median carina (extending to anterior 0.2–0.3 to posterodorsal margin of carapace); posteriormost postrostral tooth arising at about 0.2 of carapace length; dorsal margin in lateral view gently to strongly convex; supraorbital tooth moderately strong, slightly ascending, gradually tapering distally, located at level of posterior margin of orbit, not reaching tip of antennal tooth; orbital margin with distinct convexity posteriorly, base of eyestalk located between this convexity and suborbital lobe; deep V-shaped notch present below base of supraorbital tooth; suborbital lobe well developed, triangular, reaching tip of antennal tooth; anterolateral margin between antennal and pterygostomial teeth strongly sinuous with relatively shallow concavity below antennal tooth.
Abdomen ( Fig. 14 View FIGURE 14 ) dorsally rounded. Second somite with distinct transverse ridge accompanied by shallow groove on tergum. Pleura of anterior three somites broadly rounded; fourth pleuron rounded or with tiny posteroventral denticle; fifth pleuron with moderately strong posteroventral tooth. Sixth somite about 1.5–1.6 times longer than fifth somite and 1.7–1.8 times longer than high, bearing tiny posteroventral tooth; posterolateral process terminating in small tooth. Telson ( Fig. 15 View FIGURE 15 C) 1.6 times longer than sixth somite, tapering to gently convex posterior margin, bearing 4–6 dorsolateral spines on either side; posterior margin with 2 pairs of lateral spines (mesial pair longer) and 6 spiniform setulose setae ( Fig. 15 View FIGURE 15 D).
Eye ( Fig. 15 View FIGURE 15 A, B) subpyriform with stalk narrowing proximally; cornea weakly dilated, slightly wider than stalk, its maximum width approximately 0.2 of carapace length; ocellus absent.
Antennular peduncle ( Fig. 15 View FIGURE 15 A, B) reaching distal 0.2–0.3 of antennal scale. First segment distinctly longer than distal two segments combined, slightly falling short of midlength of antennal scale, dorsodistal margin armed with 2 or 3 slender teeth; stylocerite reaching or slightly overreaching distolateral margin of first peduncular segment, sharply pointed, mesial margin sinuous. Second segment about 0.4 length of first segment, with l strong dorsolateral distal tooth. Third segment less than half as long as second segment, bearing 1 small dorsodistal tooth. Lateral flagellum with thickened aesthetasc-bearing portion about 0.3 times as long as carapace.
Antenna ( Fig. 15 View FIGURE 15 A, B, E) with basicerite bearing moderately small ventrolateral tooth; carpocerite reaching midlength of antennal scale. Antennal scale 0.60 times as long as carapace and 2.5 times longer than wide; lateral margin nearly straight; distolateral tooth nearly reaching rounded distal margin of lamella.
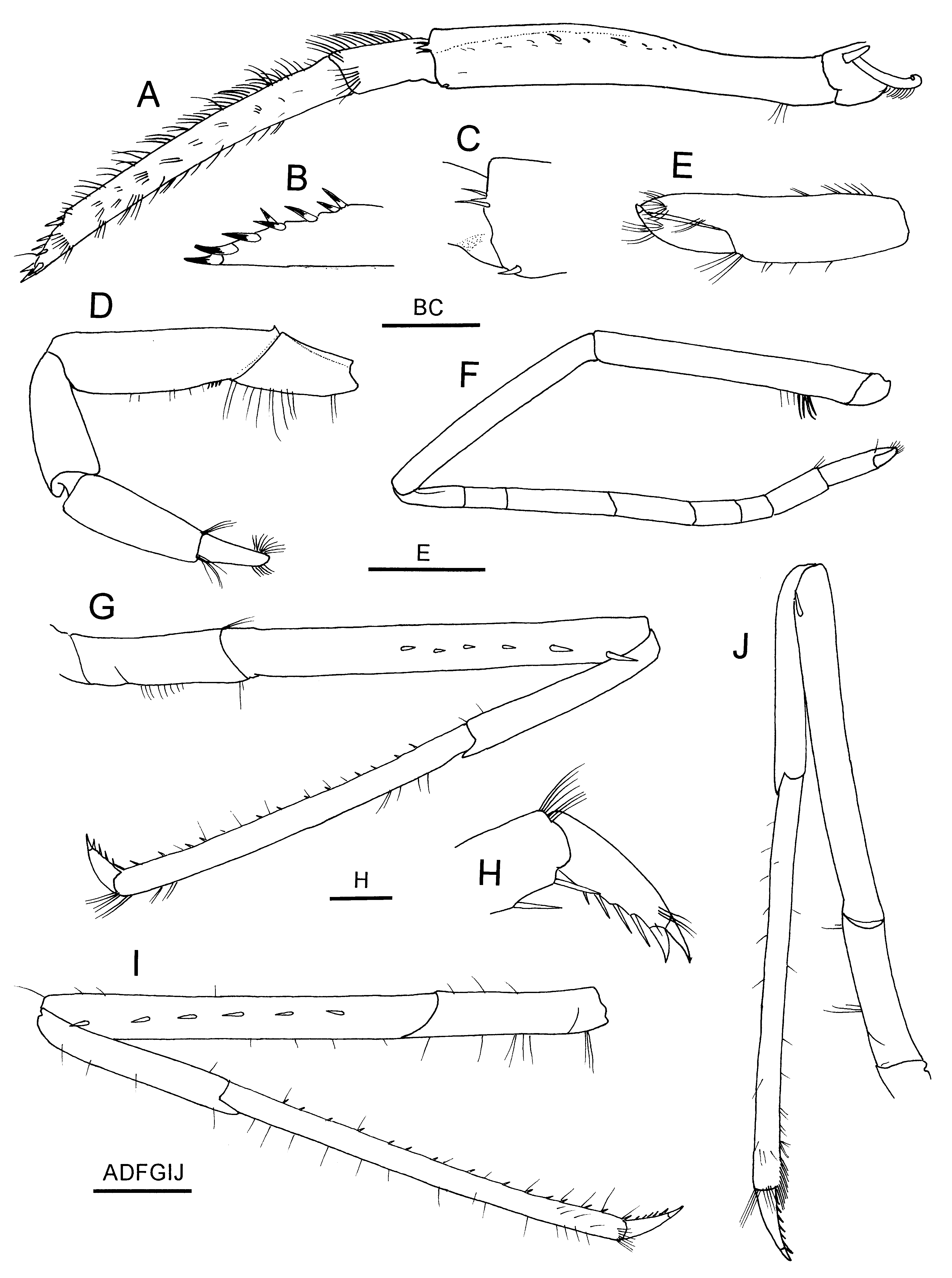
Mouthparts similar to those of other species of the genus. Third maxilliped ( Fig. 16 View FIGURE 16 A) overreaching antennal scale by half-length of ultimate segment; ultimate segment 3.6 times longer than penultimate segment, tapering distally, with short row of darkly pigmented corneous spines distomesially ( Fig. 16 View FIGURE 16 B); antepenultimate segment about 0.9 times as long as two distal segments combined, armed with 1 small tooth and 2 spiniform setae on distolateral margin and 1 spinule at ventrolateral distal angle ( Fig. 16 View FIGURE 16 C); lateral surface with row of single or paired spiniform setae on blunt ridge adjacent to dorsal margin.
Strap-like, terminally hooked epipods present on third maxilliped to third pereopod, corresponding setobranchs present on first to fourth pereopod ( Fig. 15 View FIGURE 15 F).
First pereopod ( Fig. 16 View FIGURE 16 D) moderately stout, slightly overreaching midlength of antennal scale; dactylus ( Fig. 16 View FIGURE 16 E) about 0.7 times as long as palm, terminating in 2 darkly pigmented corneous claws; fixed finger terminating in single corneous claw. Second pereopod ( Fig. 16 View FIGURE 16 F) overreaching antennal scale by about 0.4 length of carpus; carpus divided into 7 articles. Third to fifth pereopods moderately long and slender, similar in shape and subequal in length. Third pereopod ( Fig. 16 View FIGURE 16 G) overreaching antennal scale by about 0.8 length of propodus; dactylus ( Fig. 16 View FIGURE 16 H) 0.20 times as long as propodus, moderately stout (about 3.9 times longer than deep), terminating in acute, pigmented unguis, armed with 5–6 accessory spinules on flexor margin, distalmost accessory spinule subterminal, distinctly larger than others, making tip of dactylus appearing biunguiculate; carpus about 0.6 times as long as propodus; merus armed with 6 lateral spines in distal two-thirds. Fourth pereopod ( Fig. 16 View FIGURE 16 I) overreaching antennal scale by 0.4–0.5 length of propodus; merus with 4–6 lateral spines. Fifth pereopod ( Fig. 16 View FIGURE 16 J) overreaching distal margin of antennal scale by 0.2–0.3 length of propodus; merus with 1 spine at ventrolateral distal angle.
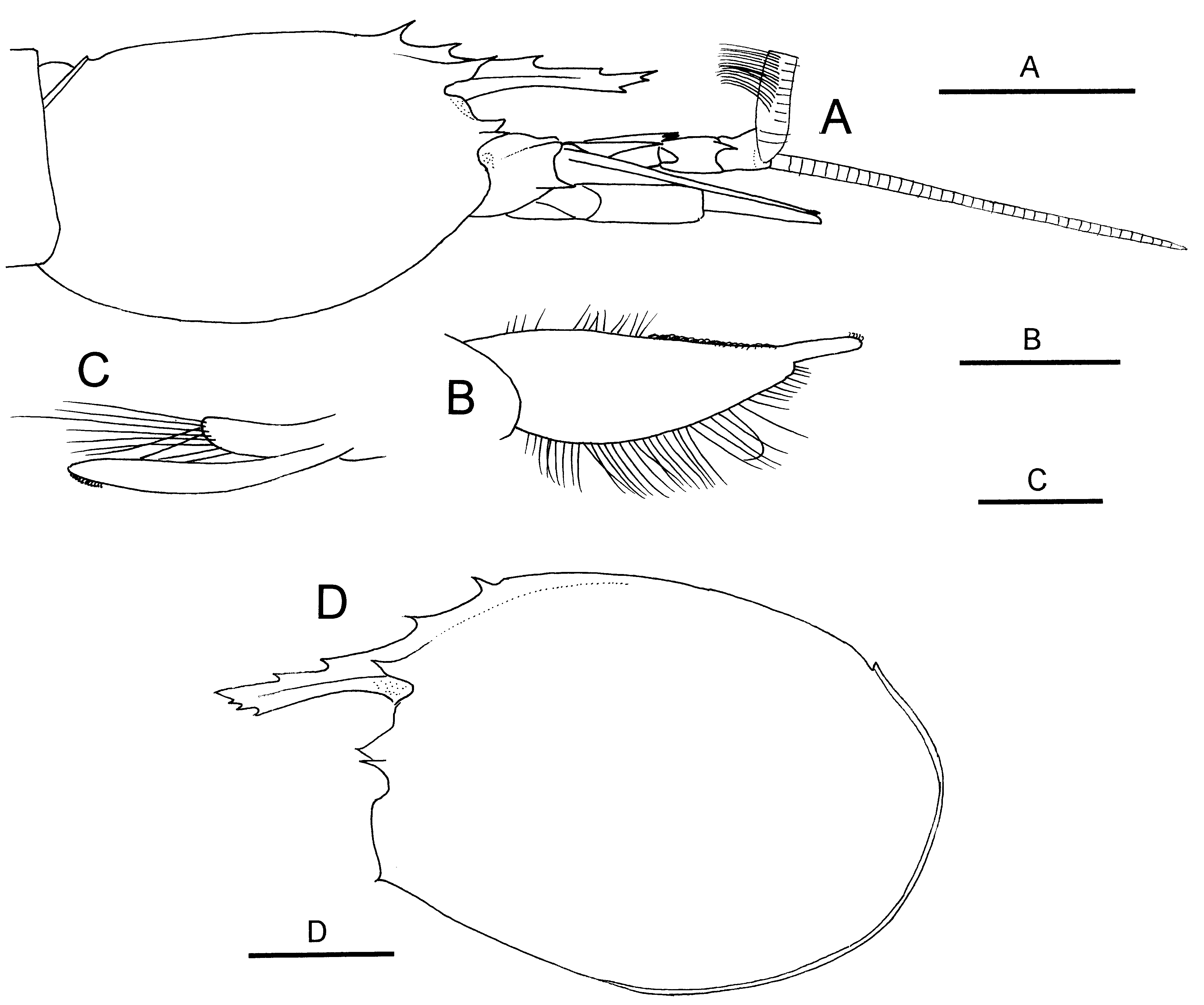
Males. Carapace ( Fig. 17 View FIGURE 17 A) with nearly straight dorsal margin in lateral view; postrostral median carina extending beyond midlength of carapace. Fourth abdominal pleuron bearing small but distinct posteroventral tooth. Antennular peduncle ( Fig. 17 View FIGURE 17 A) reaching distal 0.2 of antennal scale; outer antennular flagellum with thickened aesthetasc-bearing portion about 0.4 times as long as carapace; inner flagellum slightly longer than carapace. Endopod of first pleopod ( Fig. 17 View FIGURE 17 B) with narrow lobe just lateral to base of terminally located appendix interna. Appendix masculina on second pleopod shorter than appendix interna, bearing about 15 stiff setae ( Fig. 17 View FIGURE 17 C).
Size. Largest female cl 19.2 mm, ovigerous female cl 16.1 mm; largest male cl 9.5 mm.
Coloration in life. Not known.
Distribution. Recorded from two locations in the Southwest Pacific, Manus Basin and Lau Basin, 1512–1842 m.
Etymology. The species name, thermophilus , is the combination of the Greek, “thermos’’ (= hot), and ”philos” (= loving), in reference to the habitat of the new species.
Ecology. The associated fauna of the sites in the Lau Basin where this new species occurs, i.e., Hina Hina ( 1842 m) , Tu’i Malila ( 1890 m), and Vailili ( 1740 m), was described by Desbruyères et al. (1994). Video recordings taken during the BIOLAU cruise are available for this study. On the videos, this species is easily recognizable by the prominent eyes, as other shrimp fauna includes only alvinocaridids having reduced corneas. The videos show that individuals of Lebbeus thermophilus live among and above the molluscan beds composed of gastropods Alviniconcha hessleri Okutani & Ohta, 1998 and Ifremeria nautilei Bouchet & Warén, 1991 , and mussels Bathymodiolus brevior Cosel, Métivier, Hashimoto, 1994 , where the density of the shrimps could reach several tens of individuals/m ². The temperature of the habitat varies between 6 and 12°C ( Desbruyères et al. 1994). Some individuals were also observed at some meters of the active sites.
Remarks. The three new species described in this study all belong to the species group characterized by the possession of epipods on the anterior three pairs of pereopods, and are morphologically similar to the following 10 species: L. antarcticus , L. carinatus , L. cristatus , L. formosus , L. kuboi , L. microceros , L. polyacanthus , L. similior , L. washingtonianus , and L. wera . Shared characters include: rostrum not reaching distal margin of second segment of antennular peduncle, styliform rather than spiniform, bearing four or more dorsal teeth including postrostral teeth and more than one ventral teeth; distinct notch present inferior to base of supraorbital tooth; anterolateral margin of carapace between antennal and pterygostomial teeth strongly sinuous with deep concavity just below antennal tooth; first segment of antennular peduncle armed with more than one teeth on dorsodistal margin; and dactyli of third to fifth pereopods clearly biunguiculate. Differentiating characters among these species are summarized in Table 1.
In having an elongate antennular peduncle (reaching nearly to the distal margin of the antennal scale) with a relatively short stylocerite (not reaching the distal margin of the first segment of the antennular peduncle) and the relatively long antennal carpocerite distinctly overreaching the midlength of the antennal scale, L. pacmanus sp. nov. most closely resembles L. washingtonianus . Lebbeus pacmanus differs from L. washingtonianus in the following characters: (1) the rostrum is straight and does not reach the distal margin of the first segment of the antennular peduncle in L. pacmanus , rather than slightly curving dorsally and reaching the distal margin of the first segment of the antennular peduncle in L. washingtonianus ; (2) the dorsal rostral teeth are six, including two or three postorbital in L. pacmanus , whereas four or five including two postrostral in L. washingtonianus ; (3) the distolateral tooth of the antennal scale reaches the distal margin of the lamella in L. pacmanus , but falling short of it in L. washingtonianus ; and (4) the first pereopod overreaches the distal margin of the antennal scale in L. pacmanus , rather than just reaching it in L. washingtonianus . In the relatively short antennular stylocerite, L. antarcticus and L. similior are also similar to L. pacmanus sp. nov. The former two species differ from L. pacmanus in the shorter antennular peduncle (reaching or falling short of the base of distolateral tooth of the antennal scale versus nearly reaching the distal margin of lamella of antennal scale) and the shorter distolateral tooth of the antennal scale (distinctly falling short of the lamella versus reaching it). Furthermore, L. antarcticus is distinguished from L. pacmanus in the shorter third maxilliped (slightly overreaching the distal margin of lamella of the antennal scale versus overreaching it by the 0.7–0.9 length of the ultimate segment) and more numerous meral spines on the third and fourth pereopods (eight versus six for the third pereopod, four versus six for the fourth pereopod). Lebbeus similior is further separated from L. pacmanus by the fewer dorsolateral spines on the telson (five to seven versus four).
Among the species characterized by a relatively long antennular stylocerite (reaching or slightly overreaching the dorsodistal margin of the first segment of the antennular peduncle), L. shinkaiae sp. nov. is most similar to L. polyacanthus and L. wera in having relatively numerous dorsal rostral teeth (six or more versus less than six). However, this new species is characteristic in having the supraorbital tooth arising anterior to the rostral base and the presence of thin plumose setae in addition to the five or six spiniform setulose setae on the posterior margin of the telson. In the latter three species, the supraorbital tooth arises at the level of the rostral base or slightly posterior to it; and the posterior margin of the telson lacks thin plumose setae. Lebbeus polyacanthus is further distinguished from L. shinkaiae by the more posteriorly arising posteriormost postrostral tooth on the carapace (at 0.1–0.2 of the carapace length versus about 0.4), the constant presence of a posteroventral tooth on the fourth abdominal pleuron, much more numerous meral spines on the third to fifth pereopods (see Table 1 for the precise counts). Lebbeus wera differs from L. shinkaiae in the more posteriorly arising posteriormost postrostral tooth (at 0.3 of the carapace length versus 0.1–0.2).
Lebbeus thermophilus sp. nov. is very similar to L. carinatus , L. cristatus , L. formosanus , L. kuboi , and L. microceros in having relatively few dorsal rostral teeth (five or less) and the stylocerite reaching or slightly overreaching the dorsodistal margin of the first segment of the antennular peduncle. Lebbeus carinatus can be distinguished from L. thermophilus by the shorter third maxilliped (reaching to the distal margin of the antennal scale versus overreaching it by about 0.5 length of the ultimate segment). Lebbeus cristatus is separated from L. thermophilus by the fewer meral spines on the third and fourth pereopods (two or three versus five or six for the third pereopod, four to six versus one for fourth pereopod). Lebbeus formosanus can be separated from L. thermophilus by fewer dorsolateral spines on the telson (three or four versus four to six) and the smaller body size ( 13.7 mm in cl versus 19.2 mm). Lebbeus kuboi differs from L. thermophilus in the more elongate antennular peduncles (reaching or slightly overreaching the distal margin of the antennal scale versus far falling short of it) with a greatly elongate outer flagellum in the male and a larger body size ( 23.5 mm in cl versus 19.2 mm); furthermore, in L. kuboi , the development of epipod on the third pereopod shows individual variation (Komai et al. 2004), though it is constantly well-developed in L. thermophilus . Lebbeus microceros is easily distinguished from L. thermophilus in the different armature of the posterior margin of the telson (two spines versus three to five setulose spiniform setae) and the presence of a strong, curved distolateral tooth on the antepenultimate segment of the third maxilliped.
Wicksten & Hendrickx (1992) recorded Lebbeus washingtonianus from the Guaymas Basin, Mexico, where there is hydrothermal activity, but Martin & Haney (2005) noted that it was not known if specimens were found in association with such systems in the Guaymas Basin. Furthermore, the identification of the specimens used by Wicksten & Hendrickx (1992) need to be verified, concerning the presence of many species morphologically similar to L. washingtonianus . The previous records of L. washingtonianus from hydrothermal vents on the Okinawa Trough ( Kikuchi & Ohta 1995; Fujikura et al. 1995; Hashimoto 1997; Watabe & Miyake 2000) are here all referred to L. shinkaiae sp. nov. The unidentified species reported from the Manus and/or Lau basins ( Desbruyères et al. 1994; Komai & Segonzac 2004; Martin & Haney 2005; Komai & Collins 2009) is here described as new, L. thermophilus .
The present data suggest that the five species treated in this study are all vent-endemic. As well as the other species in the genus, each species is restricted to a rather narrow geographical area, as summarized in Table 1. It is remarkable that Lebbeus pacmanus sp. nov. and L. thermophilus sp. nov. sympatrically occur in the Manus Basin, though the latter species also occurs in the Lau Basin. Additionally, Martin & Haney (2005) reported the occurrence of Lebbeus bidentatus , originally described from off Chile at depth of 1680 m ( Zarenkov 1976), from near a vent field on the southern portion of the EPR. Komai & Collins (2009) described a new species, L. manus , from a hydrothermal vent on the Manus Basin. In total, seven species are now known from hydrothermal vents.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Caridea |
|
Family |
|
|
Genus |
Lebbeus thermophilus
| Komai, Tomoyuki, Tsuchida, Shinji & Segonzac, Michel 2012 |