
Miniopterus ambohitrensis, Goodman, Steven M., Ramasindrazana, Beza, Naughton, Kate M. & Appleton, Belinda, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3936.4.4 |
|
publication LSID |
lsid:zoobank.org:pub:26655EB2-0352-4D4F-BBD6-44F89D05DC41 |
|
DOI |
https://doi.org/10.5281/zenodo.3860414 |
|
persistent identifier |
https://treatment.plazi.org/id/03F41D33-FF89-FF84-7DE3-F89EF7E6FA8A |
|
treatment provided by |
Plazi |
|
scientific name |
Miniopterus ambohitrensis |
| status |
sp. nov. |
Miniopterus ambohitrensis View in CoL sp. nov.
Montagne d’Ambre long-fingered bat Minioptère de Montagne d’Ambre Figures 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5
Synonyms
Miniopterus manavi Peterson, Eger & Mitchell, 1995 View in CoL , in part Miniopterus aelleni Goodman, Bradman, Maminirina, Ryan, Christidis & Appleton, 2008 View in CoL , in part Miniopterus aelleni Goodman, Maminirina, Weyeneth, Bradman, Christidis, Ruedi & Appleton, 2009 View in CoL , in part Miniopterus aelleni Goodman, 2011 View in CoL , in part
Miniopterus cf. egeri Goodman, Ramasindrazana, Maminirina, Schoeman & Appleton, 2011 View in CoL Miniopterus View in CoL “ aelleni View in CoL B” Christidis, Goodman, Naughton & Appleton, 2014
Holotype. FMNH 202450, male, testes measuring 7 x 3 mm, partially convoluted epididymides, body preserved in 12% formalin and subsequently transferred to 70% ethanol, and skull removed and cleaned by Dermestidae beetles. The skull is in fine condition, with a full adult dentition, and the basisphenoid sutures fused. Original field number Steven M. Goodman ( SMG) 16169. The holotype was employed in the morphological and molecular analyses.
Type locality. Madagascar: Province d’Antsiranana, Parc National de la Montagne d’Ambre, 5.5 km SW Joffreville (Ambohitra), Station Forestière des Roussettes, 12°31’37.3”S, 49°10’19.1”E, 1000 m. Captured on 9 May 2009 in a harp trap placed along a foot path leading from the station to the Petite Cascade and in slightly disturbed montane forest.
Referred specimens used in molecular analysis. Madagascar: Province d’Antananarivo, Réserve Spéciale d’Ambohitantely, Grotte des Chauves-souris, 18°10.274'S, 47°16.898'E, 1340 m ( FMNH 194169, UADBA 43172, 43176, 43178, 43180–181). Province d’Antsiranana, Parc National de la Montagne d’Ambre, 5.5 km SW Joffreville (Ambohitra), Station Forestière des Roussettes, 12°31’37.3”S, 49°10’19.1”E, 1000 m (type locality) ( FMNH 202449, 202451–453, UADBA 31950–951, 31954, 48628–635); Parc National de Marojejy, 11.5 km SE Doany, 14.42667°S, 49.6083°E, 810 m ( FMNH 172600); Région Sofia, Bemanevika Forest, Peregrine Fund Camp, 14.34836°S, 48.57981°E, 1570 m ( FMNH 226018). See Appendix for a listing of specimens referred to this species based on morphology.
Etymology. The name ambohitrensis is derived from the geographical name of the type locality, which in Malagasy is Ambohitra and in French is Montagne d’Ambre. The nearby village of Joffreville is also referred to as Ambohitra. In the Malagasy language, the root word of Ambohitra is vohitra meaning mountain or highlands, giving an ecological context to the specific epithet of this bat, which occurs at higher elevations.
Diagnosis. A moderately small species of Miniopterus with mixed medium and dark brown colored pelage; the pectoral area and head often with varying amounts of rufous-brown fur ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , left). The holotype has a forearm length of 40 mm and based on sequenced individuals of M. ambohitrensis this measurement ranges from 37–42 mm (mean= 39.6 mm). Tragus moderately long, 6–7 mm in length, slightly broader at the base, with a slight constriction along the mid-shaft, the distal portion medially deflected, and tip slightly rounded ( Fig. 3 View FIGURE 3 , right). Cranial features include a linear palate and slightly expanded rostrum, nasal depression forming a central basin slightly more than the proximal one-half of the nasals, palatal emargination with open “v-shape” notch, and pronounced lambdoid crest ( Fig. 4 View FIGURE 4 ).
Description. Based on molecular characters, animals herein referred to M. ambohitrensis form a distinct clade and are sister to M. aelleni (see below); hereafter all references to this latter taxon are in the context sensu stricto. We have directly compared the holotypes of M. aelleni (FMNH 173067) and M. ambohitrensis (FMNH 202450) for external and cranio-dental characters.
External characters. A moderately small Miniopterus with a tail length about 40% of total length ( Table 2 View TABLE 2 ). In the holotype, which was preserved in fluid in mid-2009, the dorsal and ventral pelage is slightly long, distinctly dense, and a mixture of medium brown interspersed with a distinctly lighter brown and medium gray colored fur. In other individuals that have been genotyped to this species, the pectoral area and head often with moderate to extensive areas of rufous-brown fur ( Fig. 2 View FIGURE 2 ), which can also be variable in color intensity. Across the type series, the wing membrane and uropatagium are dark brownish-black, largely naked, and show no noticeable change in coloration across their surface area. With the exception of the rufous-brown areas of pelage, this generally coloration pattern is similar to M. aelleni .
The ear length of the M. ambohitrensis is 11 mm, and the range for the 24 specimens referred to this species is 10–12 mm (average 10.8 mm), which is basically identical with the average value of 10.8 mm for M. aelleni ( Table 2 View TABLE 2 ). The tragus length in the holotype of M. ambohitrensis is 6 mm and this species shows little variation with all animals falling with the range 6–7 mm (average 6.3 mm). In comparison, the tragus length in M. aelleni ranges from 5–6 mm (average 5.7 mm) and the difference between these two species is statistically significant (t=3.99, df=37, P =0.0003).
The tragus shape of M. ambohitrensis is moderately long, slightly broader at the base, with a lateral flange reduced distally, and towards the tip becomes medially deflected and slightly rounded ( Fig. 3 View FIGURE 3 , left). In comparison, the tragus of M. aelleni is slightly shorter, similarly broad at the base, and towards the distal tip, it tapers medially, and forms a blunt rounded tip; lateral flange not pronounced ( Fig. 3 View FIGURE 3 , right; Goodman et al. 2009a).
With the exception of ear length, the other four external measurements indicate that M. ambohitrensis is larger than M. aelleni ( Table 2 View TABLE 2 ). For example, the average forearm length in M. ambohitrensis is 39.6 mm (range 37–42 mm) and in M. aelleni is 37.1 mm (ranges 35–38 mm); the two taxa show little overlap in this measurement and this difference is statistically significant (t=7.82, df=37, P <0.0001). One external measurement that captures the diminutive size of M. aelleni with respect to M. ambohitrensis is total length with average measurements in the former of 90.0 mm (range 87–93 mm) and in the latter of 97.0 mm (range 93–102 mm); these measurements show virtually no overlap and are statistically significant (t=10.59, df=36, P <0.0001). Proportionately, M. aelleni has a slightly longer tail than M. ambohitrensis with the ratio of total length/tail length being 46.3% and 44.9%, respectively.
Total Tail Hindfoot Tragus Ear Forearm Body length length length length length length mass. ae̸̸eni sensu latο samples.
Cranio-dental characters. The skull of M. ambohitrensis has a relatively short rostrum, slightly bulbous braincase, and not notably constricted at the level of the interorbitals ( Fig. 4 View FIGURE 4 ). In dorsal view, the rostrum is slightly bulbous, particularly the nasals, which show some expansion in their medial portions, and a slight constriction in the postorbital region. Frontals somewhat rounded with prominent sagittal crest passing distally to the parietal and, in lateral view, with a flattening or depression in the parietal region ( Fig. 5 View FIGURE 5 ). Lambdoid crest is prominent and continuous to the paraoccipital process. From anterior view, the palatal emargination forms an open “v-shape” notch ( Fig. 4 View FIGURE 4 ). Medial portion of palate is flat with the lateral portions, along the lingual side of the toothrow, forming two elongated and parallel surfaces. The posterior palatal spine is relatively long and thin. The central nasal sulcus is relatively broad, forming a distinct basin, and spanning in most specimens slightly more than the proximal one-half of the nasals.
With the exception of palatal length, all of the mean measurements of cranial variables are notably greater in M. ambohitrensis than in M. aelleni and all of these differences are statistically significant ( Table 3 View TABLE 3 ). The greatest skull length in M. ambohitrensis is on average 14.9 mm (range 14.5–15.3 mm) and in M. aelleni on average 14.2 mm (range 13.5–14.3 mm); hence, there is no overlap between the two taxa.
On the basis of cranial morphology, a number of characters distinguish M. ambohitrensis relative to M. aelleni ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 ): braincase notably more bulbous, rostrum less linear in shape, nasals absolutely and proportionately broader and longer, nasal sulcus broader and deeper, basin in nasal sulcus shorter and slightly more than one-half the nasal length, frontals more rounded and with slight depression in the parietal region, sagittal crest generally more prominent and connecting to parietal crest, lambdoid crest notably prominent and continuous to paraoccipital process, palate broader, anterior portion of palatal emargination more open forming a broad “v-shaped” notch, posterior palatal spine slightly more prominent and elongated, and palate proportionately broader.
Dental formula I 2 /3 C 1/1 P 2/3 M 3/3, comprising the adult dentition of 36 teeth. As is evident in figures 4 and 5, the first upper premolar (P2) is notably reduced in size, with less complex cusp morphology than the second upper premolar (P3), all characteristics of the genus Miniopterus ( Koopman 1994) . P2 approximately one-third and P3 about one-half the length of the upper canine.
Without exception, the mean measurements of the four dental variables are greater in M. ambohitrensis than in M. aelleni ; all of these differences are statistically significant ( Table 4 View TABLE 4 ). The length of the complete cranial toothrow (I1 -M3) in M. ambohitrensis is on average 6.8 mm (range 6.5–6.9 mm) and in M. aelleni is on average 6.4 mm (range 6.1–6.5 mm); hence, there is little overlap between the two taxa and the differences are statistically significant. The same general pattern holds for the complete canine-molar toothrow (C-M3). The measurements for these two species across two portions of the upper toothrow, width across upper canines (C1-C1) and width across 3rd upper molars (M3-M3), show no overlap. In general, the dentition of M. ambohitrensis is absolutely and proportionately heavier than in M. aelleni ( Fig. 5 View FIGURE 5 ). This is, for example, evident in the upper and lower canines, as well as the first upper premolar (P2) and second upper premolar (P3).
Bioacoustics. The echolocation calls of Miniopterus ambohitrensis and M. aelleni recorded within a flight cage are characteristic of the genus with low duty cycle calls containing a broad FM component ( Fig. 6 View FIGURE 6 ). Based on the recorded calls of sequenced individuals of M. ambohitrensis and different animals referred to M. aelleni , no statistically significant difference was found for any of the measured echolocation variables (MANOVA: F5,10=1.857, P =0.189, Table 5 View TABLE 5 ).
Molecular phylogenetics and phylogeography. The same topology was recovered by both the maximum likelihood and Bayesian analyses ( Fig. 7 View FIGURE 7 ). Miniopterus ambohitrensis sp. nov. and M. aelleni were reciprocally monophyletic and united as sister species (posterior probability 1.00, bootstrap support 99). Miniopterus brachytragos was recovered as sister to this species pair with full support (1.00/100). All taxa included in the analysis were found to be reciprocally monophyletic with respect to one another and all terminal nodes showed a high level of support, although M. aelleni only showed node support of 74 in the ML analysis. This is not considered problematic due to the Bayesian posterior probability at that node (1.00) and the high level of resolution throughout the remainder of the tree.
Pairwise mean genetic distance between taxa is presented in Table 6 View TABLE 6 . Miniopterus ambohitrensis differs from M. aelleni by 3.3% K2P across the 725 bp cytochrome b sequences sampled. The intraspecific distances within M. ambohitrensis and M. aelleni were 0.8% and 1.1%, respectively.
Aspects of the natural history. The type locality of Miniopterus ambohitrensis is in close proximity to the Station des Roussettes in the Parc National de la Montagne d’Ambre in extreme northern Madagascar (Fig. 1). This massif, which rises out of the coastal plain, is volcanic in origin, the summit at 1476 m, and locally referred to as Ambohitra. Considerable volcanic activity took place on the mountain during the Upper Pliocene and Quaternary ( Brenon 1972; Karche 1972 in Rossi 1974), with evidence of large scale eruptions until about 8,000 years ago ( Battistini 1965).
An altitudinal transect of precipitation patterns on the massif shows pronounced variation in annual rainfall: 3.9 m /year at the Station des Roussettes, 2.2 m /year at Joffreville (700 m and 5.5 km from the station), and 1.3 m / year at Sakaramy (380 m and 15 km direct distance from the station) ( Donque 1975). Vegetational variation along these slopes is directly correlated with orography: the zone from the mountain base to approximately 800 m is composed of dry deciduous or at the higher reaches of this zone of transitional dry deciduous-humid forest, abruptly at around 900 m there is a shift to humid forest, and in the last few hundred meters of elevation to the summit there is increasingly heavier epiphytic loads on woody vegetation and exposed rocks, as well as a diminution of tree stature.
All of the individual bats from Montagne d’Ambre identified as M. ambohitrensis were captured in the park and more specifically in the immediate vicinity of the Station des Roussettes, approximately at 1000 m (Appendix). Many of the specimens allocated to this species were captured along a footpath at the edge of the forest between the station and the Petite Cascade a few hundred meters away. Further, this species is known to utilize a shallow cave at the base of the Petite Cascade as a day roost site.
The vegetation types at the other sites M. ambohitrensis has been documented are all mid-elevation or montane humid forest (Fig. 1). It has been captured at elevations as low as 810 m on the northwestern slopes of Marojejy and as high as 1570 m on the southern slopes of the Tsaratanana Massif (Bemanevika). In notable contrast, the sister species, M. aelleni , is restricted to dry forest formations, including tsingy habitat associated with limestone and karst landscapes, such as the Ankarana Massif, where the holotype of M. aelleni (FMNH 173067) was collected. This site is about 44 km direct distance from the type locality of M. ambohitrensis on Montagne d’Ambre. On the basis of current information, these two sister taxa utilize notably different biotopes: M. ambohitrensis in upland areas with mesic or montane forest and M. aelleni in lowland areas with dry deciduous formations.
Animals referred to M. ambohitrensis have been found on Montagne d’Ambre during the months of February to May (Appendix). Insufficient information is available about the presence of this species on this massif across the calendar year and it is not clear if the local population is at least partially migratory.
The sex ratio of captured animals from Montagne d’Ambre is nearly even. Evidence of recent reproductive activity was found in early April 2009, with one female lactating and possessing a single placental scar (UADBA 48630), and several females with large mammae and missing fur (UADBA 48632–634). A male taken during this same period had testes measuring 4 x 3 mm and the epididymides partially convoluted (UADBA 48629) and a female with large mammae showed signs of estrous (UADBA 48628). The latter record of late seasonal breeding and apparent post-partum estrous may indicate that after mating there is a period of reproductive dormancy, as known from some African members of this genus (van der Merwe 1979; Bernard 1980; Onyango et al. 1995).
As presented in more detail in the discussion, several different phylogroups of small Miniopterus spp. have been identified from the Réserve Spéciale d’Ambohitantely, roosting together in the Grotte des Chauves-souris ( Christidis et al. 2014). Regardless of the visitation period, the bats handled in the cave are disproportionately composed of males: 10 January 2007 – 8♂♂, 0♀; 1–8 April 2014 – 25♂♂ (several with 5 x 4 mm testes with partially convoluted epididymides), 7♀♀ (all with small mammae and no sign of recent reproduction); and 20 November 2008 —11 ♂♂, 1♀. Of the more than 50 Miniopterus handled at this site, no clear evidence of active breeding has been found, most particularly amongst females. Different African populations of Miniopterus , such as M. natalensis , are known to display elevational migration, specifically between lowland breeding areas and montane hibernacula (van der Merwe 1975; Voigt et al. 2014). It is conceivable that the same pattern occurs on Madagascar, although direct evidence is lacking and mark-recapture studies are needed.
Inside for M. ambohitrensis the Grotte des Chauves-souris, bat composition and abundance show considerable variation between visits. Miniopterus spp. were found roosting in syntopy with Myotis goudoti and in one occasion with Triaenops menamena ( Ramasindrazana & Goodman 2014) . Following the different capture sessions within this cave noted above, the greatest number of Miniopterus was observed in November.
Distribution and conservation status. The past years have seen a considerable augmentation of information on the Madagascar chiropteran fauna, particularly aspects concerning taxonomy and faunistics, which have been summarized in two recent publications (Goodman 2011; Goodman & Ramasindrazana 2013). Herein, we describe Miniopterus ambohitrensis , the eighth species of Miniopterus to be named from the island since 2007. This morphologically and genetically distinct species is currently known from four localities and insufficient information is available on its distribution and population numbers to approach a relatively accurate assessment of its conservation status. It is probably dependent on caves and rock shelters for day roosts and, hence, the availability of such sites is an important factor in determining its distribution.
Three of the four sites we have identified specimens of M. ambohitrensis from (Parc National de Marojejy, Parc National de la Montagne d’Ambre, and Réserve Spéciale d’Ambohitantely), are part of the existing protected areas system of Madagascar. The fourth site, Bemanevika, has been proposed as a new protected area ( Rabearivony et al. 2010). At all of these sites, considerable areas of forest remain; although, it is unclear if M. ambohitrensis is forest dependent for any aspect of its life cycle. At Ambohitantely, this species has been found roosting in a cave within the largest remaining forest block in this reserve; at Marojejy, foraging along a river in an extensive forest; at Montagne d’Ambre, flying though a disturbed corridor within forest and adjacent to large block of native montane habitat; and at Bemanevika, moving along a river channel surrounded by disturbed forest.
A polygon encompassing all of the sites M. ambohitrensis is known from and falling within the elevational range from 800 to 1600 m produces an “extent of occurrence”, as defined by the IUCN (IUCN 2012), of 15,143 km 2 (Fig. 1). As this species is known from upland areas with montane forest, natural habitats that at this time not particularly threatened by human activities ( Harper et al. 2007), and that it occurs in degraded forest formations, in the short or medium term probably it is not facing any severe threats to its continued existence. Field studies are needed to have a more thorough understanding of this species’ distribution, population status, and, in turn, produce accurate estimations of its conservation status. Given information presented in the previous section, it is possible that this species is at least partially migratory.


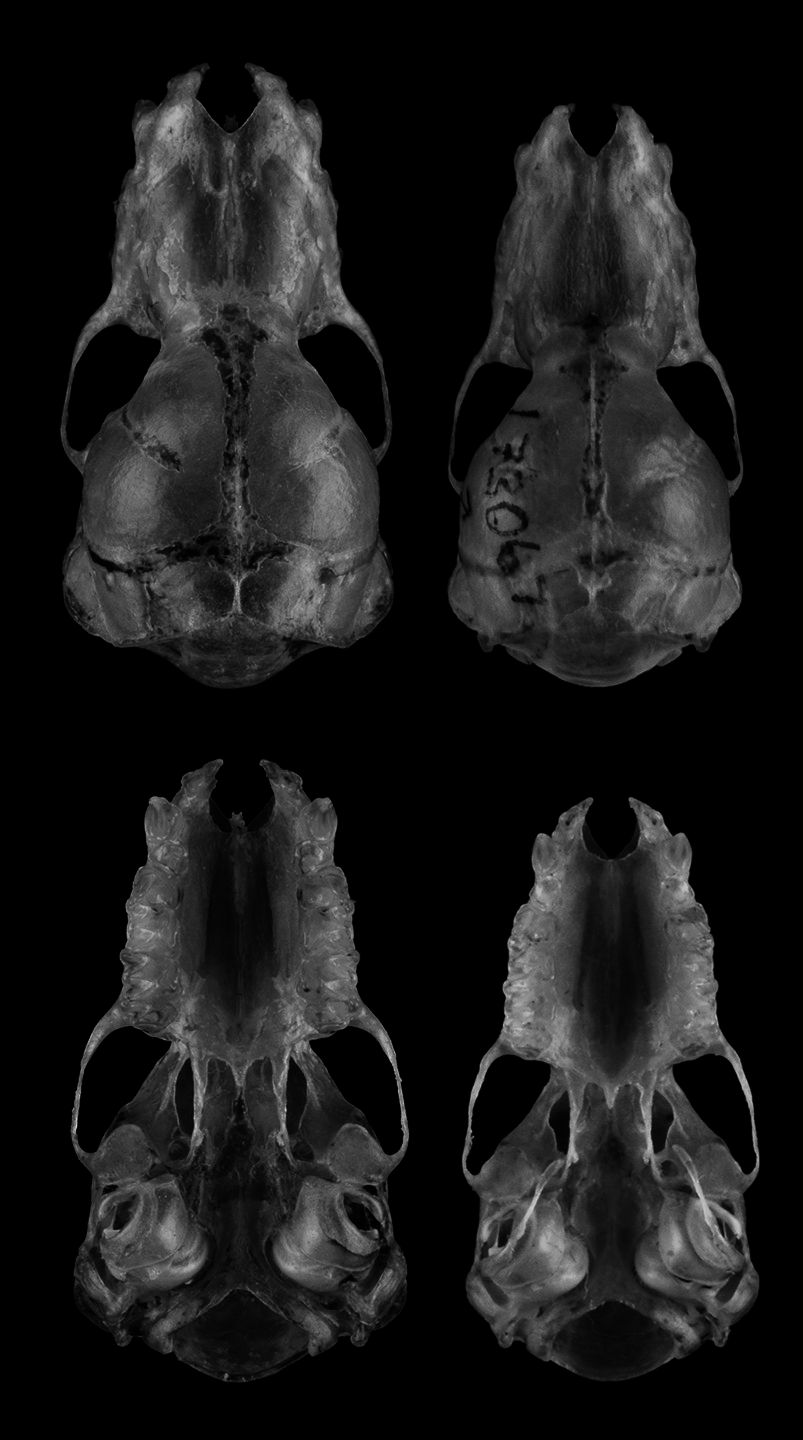



TABLE 2. External measurements (in millimeters) and mass (in grams) of adult (sexes combined) Miniopterus ambohitrensis sp. nov. and M. aelleni sensu stricto. Measurements were all taken by the same field collector (SMG) and presented as mean ± standard deviation, minimum and maximum measurements, and number of specimens. Statistical differences between M. ambohitrensis and M. aelleni based on student t-tests, n. s. = not significant.
| M. ambohitrensis Holotype ♂ FMNH 202450 | 100 | 41 | 5 | 6 | 11 40 | 7.7 |
|---|---|---|---|---|---|---|
| M. ambohitrensis | 97.0 ± 2.18 93 ‾ 102, n=23 | 43.6 ± 1.74 40 ‾ 47, n=24 | 6.1 ± 0.50 5 ‾ 7, n=24 | 6.3 ± 0.44 6 ‾ 7, n=24 | 10.8 ± 0.48 39.6 ± 0.94 10 ‾ 12, n=24 37 ‾ 42, n=24 | 6.4 ± 0.63 5.3 ‾ 7.7, n=24 |
| M. aelleni Holotype ♀ FMNH 173067 | 89 | 43 | 7 | 6 | 11 38 | 3.9 |
| M. aelleni | 90.0 ± 1.60 87–93, n=15 | 41.7 ± 1.92 39–44, n=15 | 5.7 ± 0.72 5–7, n=15 | 5.7 ± 0.49 5–6, n=15 | 10.8 ± 0.68 37.1 ±1.04 10–12, n=15 35–38, n=15 | 4.5 ± 0.49 3.7–5.4, n=15 |
| T-statistics for M. ambohitrensis compared to M. aelleni | T=10.59 P <0.0001 | T=3.48 P =0.001 | T=2.12 P =0.04 | T=3.99 P =0.0003 | T=7.82 n.s. P <0.0001 | T=9.64 P <0.0001 |
TABLE 3. Cranial measurements (in millimeters) οf hοlοtypes and adult (sexes cοmbined) Miniopterus ambohitrensis sp. nov. and M. ae ̸̸ eni sensu strictο. Measurements presented as mean ± standard deviatiοn, minimum and maximum measurements, and number οf specimens. See Materials and Methοds fοr an explanatiοn οf variable acrοnyms. Statistical differences between M. ambohitrensis and M. ae ̸̸ eni are examined based οn student t-tests. Τhe descriptive statistics presented here fοr M. ae ̸̸ eni sensu strictο are different frοm thοse in Gοοdman et a ̸ • (2009 a), as the larger M. ambohitrensis have been remοved frοm
| M. ambohitrensis Hοlοtype ♂ FMNH 202450 | GSKL 15.1 | CIL ΖΥGΟ 14.6 8.0 | PΟB 3.3 | MASΤ 7.1 | GBW 8.0 | PAL 5.5 | LW 4.4 | MAND 10.5 |
|---|---|---|---|---|---|---|---|---|
| M. ambohitrensis | 14.9 ± 0.26 14.5-15.3, n=17 | 14.3 ± 0.27 7.9 ± 0.12 13.9-14.9, 7.7-8.1, n=17 n=15 | 3.2 ± 0.13 3.0-3.4, n=17 | 7.1 ± 0.12 6.9-7.3, n=17 | 7.8 ± 0.17 7.4-8.1, n=17 | 5.5 ± 0.20 5.3-6.0, n=15 | 4.2 ± 0.13 4.0-4.4, n=17 | 10.5 ± 0.20 10.1-10.8 n=17 |
| M. ae̸̸eni ♂ Hοlοtype FMNH 173067 | 14.2 | 13.5 7.1 | 3.0 | 6.5 | 7.0 | 5.6 | 3.8 | 9.7 |
| M. ae̸̸eni | 13.9 ± 0.22 13.5-14.3, n=14 | 13.3 ± 0.24 7.2 ± 0.10 12.9-13.6, 7.0-7.3, n=13 n=11 | 3.0 ± 0.11 2.8-3.2, n=15 | 6.6 ± 0.16 6.2-6.8, n=15 | 7.1 ± 0.15 6.9-7.3, n=15 | 5.3 ± 0.17 4.9-5.6, n=13 | 3.9 ± 0.13 3.8-4.2, n=15 | 9.6 ± 0.26 8.9-9.9, n=14 |
| Τ-statistics fοr M. ambohitrensis cοmpared tο M. ae̸̸eni | Τ=10.49 P <0.0001 | Τ=10.68 Τ=16.78 P <0.0001 P <0.0001 | Τ=4.00 P <0.0001 | Τ=11.07 P <0.0001 | Τ=12.65 P <0.0001 | Τ=3.83 P =0.0007 | Τ=6.19 P <0.0001 | Τ=9.81 P <0.0001 |
TABLE 4. Dental measurements (in millimeters) of holotypes and adults (sexes combined) of Miniopterus ambohitrensis sp. nov. and M. aelleni sensu stricto. Measurements presented as mean ± standard deviation, minimum and maximum measurements, and number of specimens. See Materials and Methods for an explanation of variable acronyms. Statistical differences between M. ambohitrensis and M. aelleni are examined based on student t-tests. The descriptive statistics presented here for M. aelleni sensu stricto are different from those in Goodman et al. (2009 a), as the larger M. ambohitrensis have been removed from M. aelleni sensu lato samples.
| M. ambohitrensis Holotype ♂ FMNH 202450 | I1-M3 6.8 | C-M3 5.5 | C1-C1 4.5 | M3-M3 5.7 |
|---|---|---|---|---|
| M. ambohitrensis | 6.7 ± 0.12 6.5–6.9, n=17 | 5.5 ± 0.14 5.3–5.7, n=17 | 4.3 ± 0.13 4.0–4.5, n=17 | 5.6 ± 0.10 5.5–5.8, n=17 |
| M. aelleni Holotype ♂ FMNH 173067 | 6.4 | 5.4 | 3.7 | 5.2 |
| M. aelleni | 6.3 ± 0.12 6.1–6.5, n=15 | 5.3 ± 0.12 5.0–5.4, n=15 | 3.7 ± 0.09 3.5–3.8, n=15 | 5.2 ± 0.09 5.1–5.4, n=15 |
| T-statistics for M. ambohitrensis compared to M. aelleni | T=9.06 P <0.0001 | T=5.71 P <0.0001 | T=14.79 P <0.0001 | T=11.74 P <0.0001 |
TABLE 5. Comparison of the echolocation parameters of Miniopterus ambohitrensis sp. nov. and M. aelleni sensu stricto recorded inside a flight cage. Measurements presented as mean ± standard deviation, minimum – maximum, and number of individuals analyzed.
| Species | PF (kHz) | Fmax (kHz) | Fmin (kHz) | Dur (ms) | IPI (ms) |
|---|---|---|---|---|---|
| M. ambohitrensis | 54.3 ± 0.82 53.1–55.4 n=9 | 68.8 ± 4.44 64.0–78.2 n=9 | 49.6 ± 1.23 47.7–51.6 n=9 | 2.9 ± 0.35 2.5–3.4 n=9 | 73.4 ± 6.04 61.4–73.4 n=9 |
| M. aelleni | 53.4 ± 1.58 50.7–55.5 n=7 | 68.1 ± 5.74 62.3–79.6 n=7 | 50.0 ± 1.08 47.7–51.2 n=7 | 3.2 ± 0.82 2.2–4.8 n=7 | 78.25 ± 18.6 42.0–95.5 n=7 |
TABLE 6. Kimura 2 - Parameter (K 2 P) genetic distance between lineages of Miniopterus spp. derived from 725 bp cytochrome b sequences. Italicized values refer to K 2 P distance within the lineage based on included sequences.
| M. aelleni | M. ambohitrensis | M. brachytragos M. egeri | M. manavi | |
|---|---|---|---|---|
| M. aelleni | 0.011 | |||
| M. ambohitrensis | 0.033 | 0.008 | ||
| M. brachytragos | 0.067 | 0.072 | 0.017 | |
| M. egeri | 0.078 | 0.086 | 0.090 0.008 | |
| M. manavi | 0.089 | 0.088 | 0.081 0.043 | 0.006 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Miniopterus ambohitrensis
| Goodman, Steven M., Ramasindrazana, Beza, Naughton, Kate M. & Appleton, Belinda 2015 |
Miniopterus aelleni
| Goodman 2011 |
Miniopterus cf. egeri
| Goodman, Ramasindrazana, Maminirina, Schoeman & Appleton 2011 |
Miniopterus aelleni
| Goodman, Maminirina, Weyeneth, Bradman, Christidis, Ruedi & Appleton 2009 |
Miniopterus aelleni
| Goodman, Bradman, Maminirina, Ryan, Christidis & Appleton 2008 |
Miniopterus manavi
| Peterson, Eger & Mitchell 1995 |