
Blackburnia fulgida Liebherr, 2000: 452
|
publication ID |
https://doi.org/ 10.5281/zenodo.8159677 |
|
DOI |
https://doi.org/10.5281/zenodo.8200582 |
|
persistent identifier |
https://treatment.plazi.org/id/03F2B64D-8844-8444-92C1-62D5FDC0FAB5 |
|
treatment provided by |
Felipe |
|
scientific name |
Blackburnia fulgida Liebherr, 2000: 452 |
| status |
|
Blackburnia fulgida Liebherr, 2000: 452 , in Liebherr and Zimmerman 2000.
Diagnosis. The male specimen can be placed as a member of the Blackburnia subgenus Metromenus Sharp ( Liebherr and Zimmerman 2000: 56) based on: 1, its brachypterous condition; 2, presence of dorsolateral sulci on the basal two meso- and metatarsomeres; 3, well developed eyes; and 4, glabrous scape bearing only a single macroseta on the dorsoanterior surface. Within subgenus Metromenus , the male specimen affirms the initial species diagnosis ( Liebherr and Zimmerman 2000: 452), including: 1, upper body surface glossy, the pronotal disc smooth with only indistinct transverse lines visible within depressed portions of the cuticle; 2, basal pronotal seta present,
lateral pronotal seta absent; 3, apical lobes of metatarsomere 4 short; and 4, pronotum broad basally, MPW/BPW = 1.14 (holotype female) to 1.20 (male specimen). The eyes are moderately convex and the head narrow, resulting in ocular ratios of 1.62 for both specimens. The standardized body lengths are both 8.0 mm. For the female holotype this measurement is 0.3 mm greater than that reported in Liebherr and Zimmerman (2000: 452). The only amendment needed to the identification key in Liebherr and Zimmerman (2000: 68) is to note the presence of evident, shallowly margined isodiametric sculpticells on the elytra, those sculpticells difficult to trace in areas of reflected light. More well-developed microsculpture is commonly observed in mature versus teneral adult carabid beetles.
The male specimen is fully sclerotized ( Fig. 1A View Figure 1 ), with the elytra and pronotal margins opaque in contrast to those of the teneral female holotype ( Liebherr and Zimmerman 2000, fig. 278a).
Nonetheless, the body of this species is pale, with the dark brunneous pronotal and elytral discs contrasted with the flavous margins, and the legs, including the femora, tibiae and tarsi flavous. This pale coloration differentiates this species from the other Haleakala species, B. insociabilis (Blackburn) , which is characterized by a piceous body with glossy reflection and brunneous legs. Moreover, B. insociabilis is smaller; standardized body length 7.2–7.5 mm ( Liebherr and Zimmerman 2000, 457). In addition, the aedeagal median lobe apex of B. fulgida is narrowly rounded ( Fig. 1C View Figure 1 ), versus more broadly rounded and parallel sided in B. insociabilis ( Liebherr and Zimmerman 2000: fig. 280a).
Male genitalia. Antecostal margin of tergite VIII ( Deuve 1993) angulate distally, angulate apex slightly broader than lateral arms ( Fig. 1B View Figure 1 ); aedeagal median lobe lightly melanized, translucent in cleared specimen, lightly spiculate internal sac folds visible through wall of lobe ( Figs 1B, C View Figure 1 ); basal bulb smooth, without sagittal crest; lobe gracile, slightly broadened dorsoventrally near midlength, ventral margin straight near midlength, and ventral surface slightly depressed beyond straight portion before ventral curvature before apex ( Fig. 1C View Figure 1 ; lobe apex narrowed from edge of ostial opening to narrowly rounded, subacuminate tip).
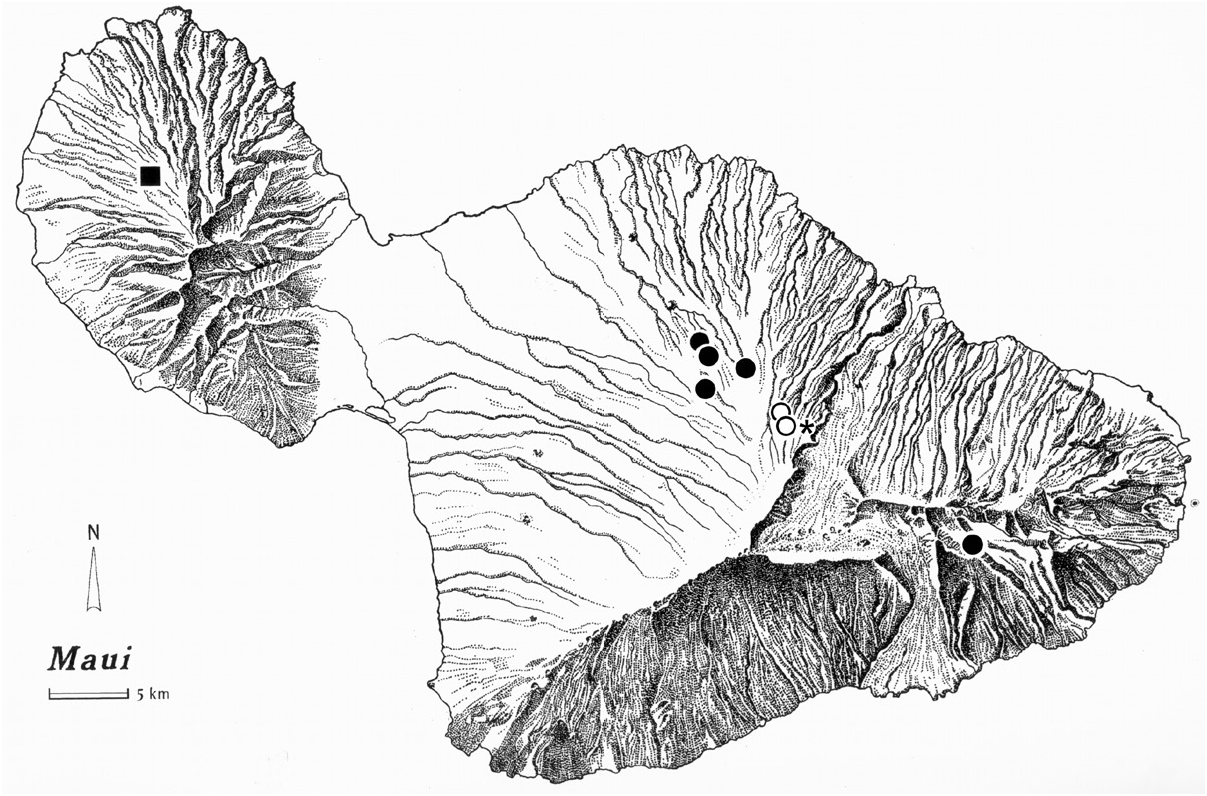
Distribution. The male specimen bears locality information: HI: Maui Is. Haleakala, Waikamoi N.C.P., 1760 m el., 20°47.21´N 156°13.82´W, 12-iii-2002, R. Takumi [Kaholoaa], pyrethrin fog mossy Ohia. The specimen was found in association with one specimen each of Blackburnia abaxoides Liebherr , B. fracta (Sharp) , and B. sphodriformis (Sharp) , and 8 specimens of B. erro (Blackburn) . The newly reported locality is only slightly downhill from the type locality ( Fig.2 View Figure 2 ).
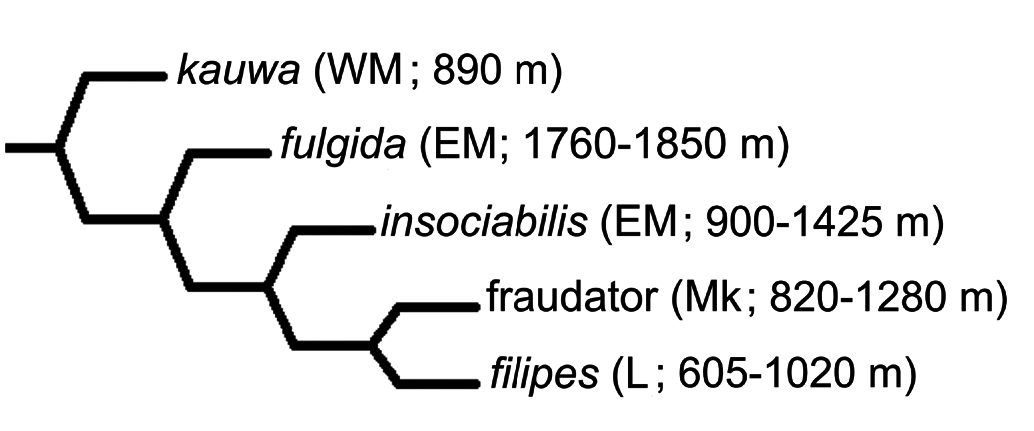
Phylogenetic placement. Cladistic analysis under parsimony using Nona running within Winclada resulted in 1727 equally parsimonious trees of 1137- step length after 5000 ratchet iterations, and 3085 trees of the same length in the 10,000 iteration analysis; sets of equally parsimonious trees of length equal to those previously reported in Liebherr and Porch (2015). Hard collapsing these trees to eliminate all trees ambiguously supported under fast or slow optimizations left 827 unambiguously optimized trees for the 5000 ratchet run, versus 1143 unambiguously optimized trees in the 10,000 ratchet analysis. Strict consensus of trees in both subanalyses collapses 26 nodes, resulting in a 1225-step consensus tree with relationships also identical to that reported by Liebherr and Porch (2015, fig. 2B). In the five-species clade ( Fig. 3 View Figure 3 ), B. fulgida is adelphotaxon to the threespecies clade ( B. insociabilis ( B. fraudator + B. filipes )). All of these species are distributed on fragments of Maui Nui, with B. fulgida and B. insociabilis both distributed on Haleakala, East Maui ( Fig. 3 View Figure 3 ). The B. insociabilis locality records are widespread from Olinda to Kipahulu Valley, at elevations from 900–1425 m ( Liebherr and Zimmerman 2000), whereas the two B. fulgida localities range from 1760–1850 m elevation. Within that elevational range, one specimen was collected from underneath a loose bark flap of an Acacia koa tree, and the second from a mossy Ohia branch; both situations within the Koa/Ohia Montane Wet Forest ( Gagné and Cuddihy 1990). Thus based on available evidence, sympatry between B. insociabilis and B. fulgida appears to be allopatrically dissected by elevation, with B. fulgida found near the upper reaches of the montane forest approaching treeline. Elevational occurrences of the other three species in this clade match that of B. insociabilis , with these species occurring within cloud forest habitats at elevations between 605 and 1280 m. That B. fulgida can occupy high elevation forest well above 1700 m elevation is made possible by the greater stature of Haleakala. That this species is not known to extend to lower elevation forest conversely suggests that the ecological preference of B. fulgida centers on the higher elevation Koa/Ohia Montane Wet Forest that can be characterized by lower levels of rainfall, and also deeper, more well-aerated soils housing symbiotic mycorrhizal Bradyrhizobium Jordan bacteria that support Acacia koa ( Nakao and Kitayama 1996) . Haleakala is the only mountain of Maui Nui tall enough to house such high-elevation mixed Koa/ Ohia forest. Both Lanai and Molokai lack any habitats of this elevation, and Puu Kukui, West Maui attains 1760 m elevation, but has only limited tracts of low-stature Ohia Montane Wet Forest near its summit ( Gagné and Cuddihy 1990). Thus B. fulgida stands out as a high-elevation specialist among species of this clade.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Blackburnia fulgida Liebherr, 2000: 452
| Liebherr, James K. & Kahoolaa, Raina L. 2020 |
Blackburnia fulgida
| Liebherr, J. K. & E. C. Zimmerman 2000: 452 |