
Tetanurae, Gauthier, 1986
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2022v44a12 |
|
publication LSID |
urn:lsid:zoobank.org:pub:231E3A73-1800-4559-A995-013482DEA623 |
|
DOI |
https://doi.org/10.5281/zenodo.6399029 |
|
persistent identifier |
https://treatment.plazi.org/id/03F29C74-FFE6-960F-D42E-6AD5FF2A9906 |
|
treatment provided by |
Felipe |
|
scientific name |
Tetanurae |
| status |
|
Tetanurae indet.
AXIAL SKELETON
Dorsal vertebrae
Material. dorsal vertebrae B4( Fig.14 View FIG )and MPV 2020.1.7 ( Fig.15A, B View FIG ) and posterior dorsal vertebra MPV V869 ( Fig. 15C, D View FIG ).
Stratigraphy. Oxfordian or Callovian marls ( MPV 2020.1.7 and B4). Lower Oxfordian marls according to the A. gregareum shells in the matrix ( MPV V869).
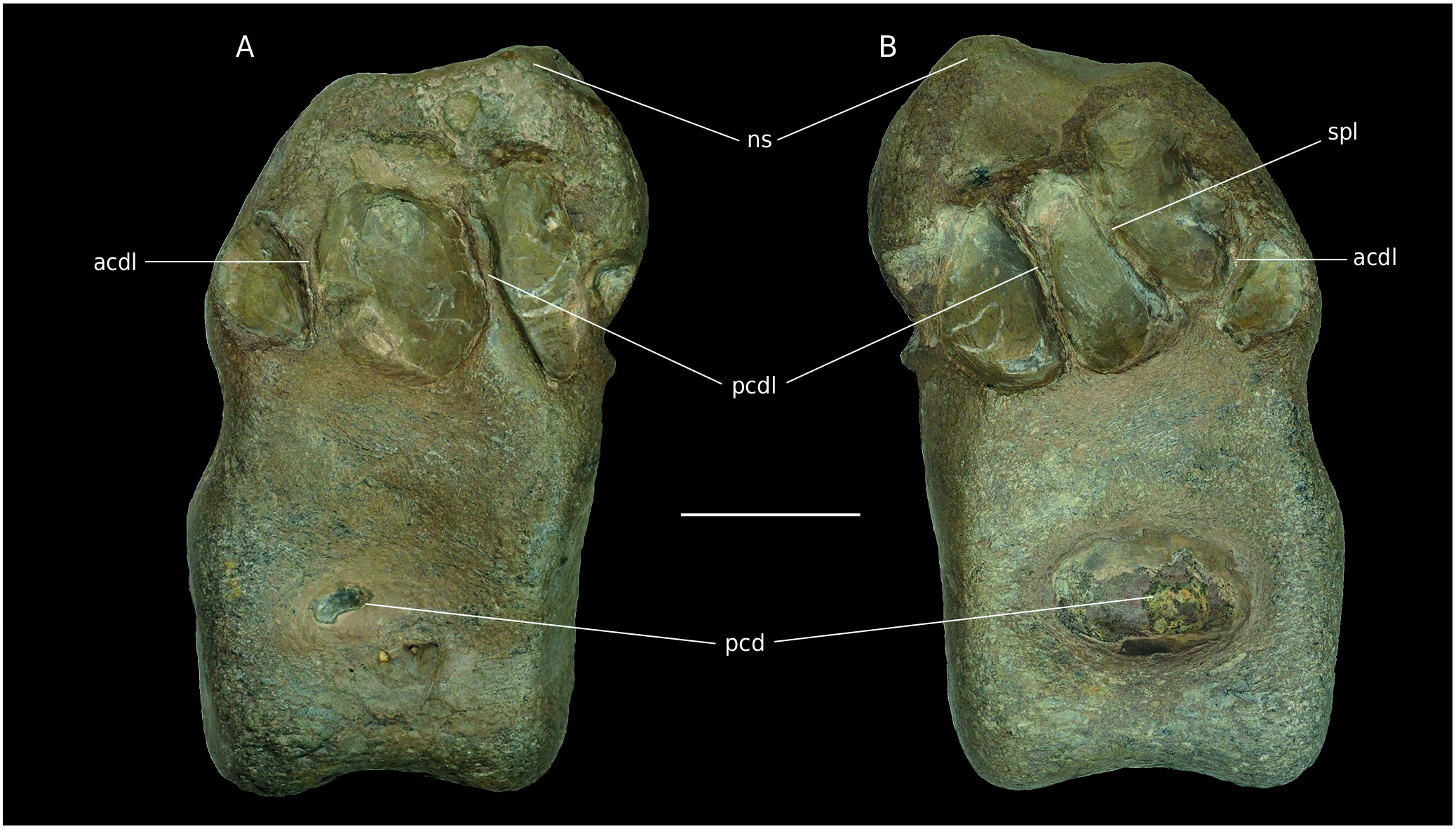
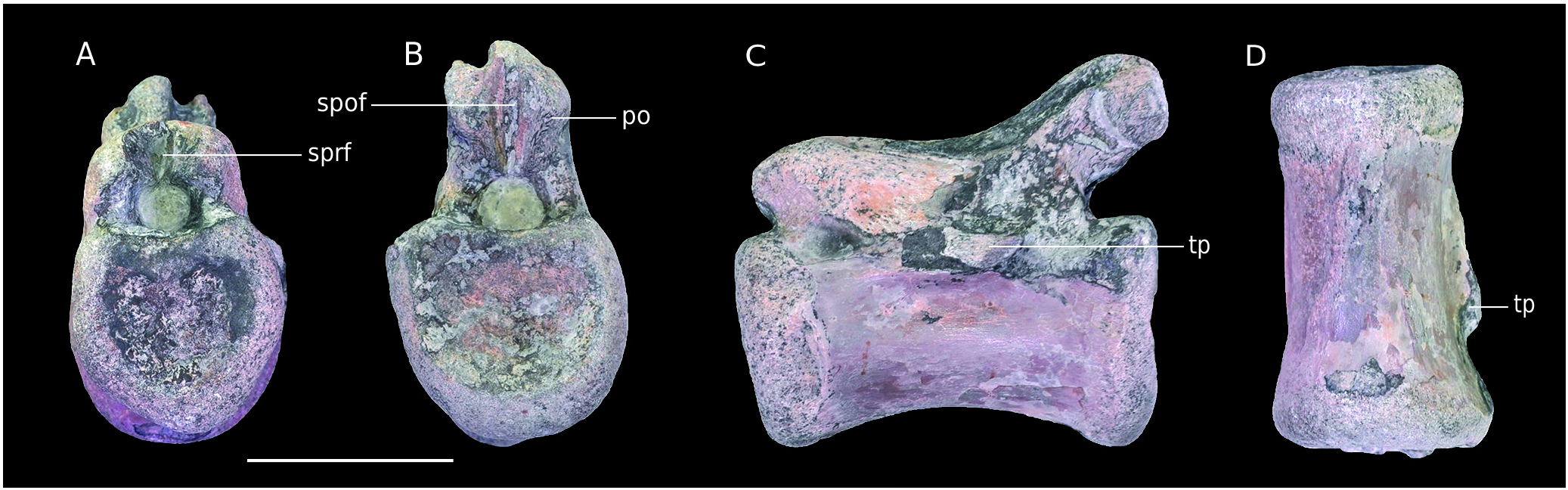
B4. The vertebra is so eroded and water-worn that it does not appear to be a bone at first sight. Despite the strong erosion, the neural arch is still preserved and attached to the centrum. All of the apophyses are missing and only the base of the neural spine is visible. Articular facets are not preserved. This preservation state is remarkable and differs from other Vaches Noires remains studied here. It is the result of a long time period on the foreshore.
The bottom of a pleurocentral depression is still visible on each lateral side. According to the strong erosion, these should have been quite deep. The neural canal is anteriorly and posteriorly visible as a rounded depression. Remains of the hyposphene are present, above a triangular depression and allow the orientation of the specimen. Oddly enough, the bone laminae of the neural arch are clearly visible ( Fig. 14 View FIG ). Despite the absence of the apophyses, anterior and posterior centrodiapophyseal as well as centropostzygapophyseal laminae are observable and frame their corresponding fossae ( Wilson et al. 2011) Another peculiarity of the specimen is the asymmetry of these laminae. Indeed, in right lateral view, an additional lamina is visible in the centre of what should be the centrodiapophyseal fossa ( Fig. 14B View FIG ). The lamina issues from the same base as the anterior centrodiapophyseal lamina but its end is posteriorly oriented. This additional structure comes with a different orientation of the dorsal end of the posterior centrodiapophyseal lamina on each side of the vertebra. Indeed, the latter is anteriorly oriented on the right side and posteriorly oriented on the left side. The dorsal margin of the spinopostzygapophyseal fossa is anteriorly oriented and is elliptical, as well as the spinoprezygapophyseal fossa.
Because parapophyses are absent and the specimen is very eroded, a clear positioning of the vertebra is impossible. According to the phylogeny of Carrano et al. (2012), the anterior orientation of the neural spine on dorsal vertebrae is a synapomorphy of Allosauroidea . However, considering the poorly preservation of the specimen a taxonomic assignment above Tetanurae will not be attempted. Moreover, the preserved part of the neural spine is too incomplete to discern a real character state and this orientation could be artificial.
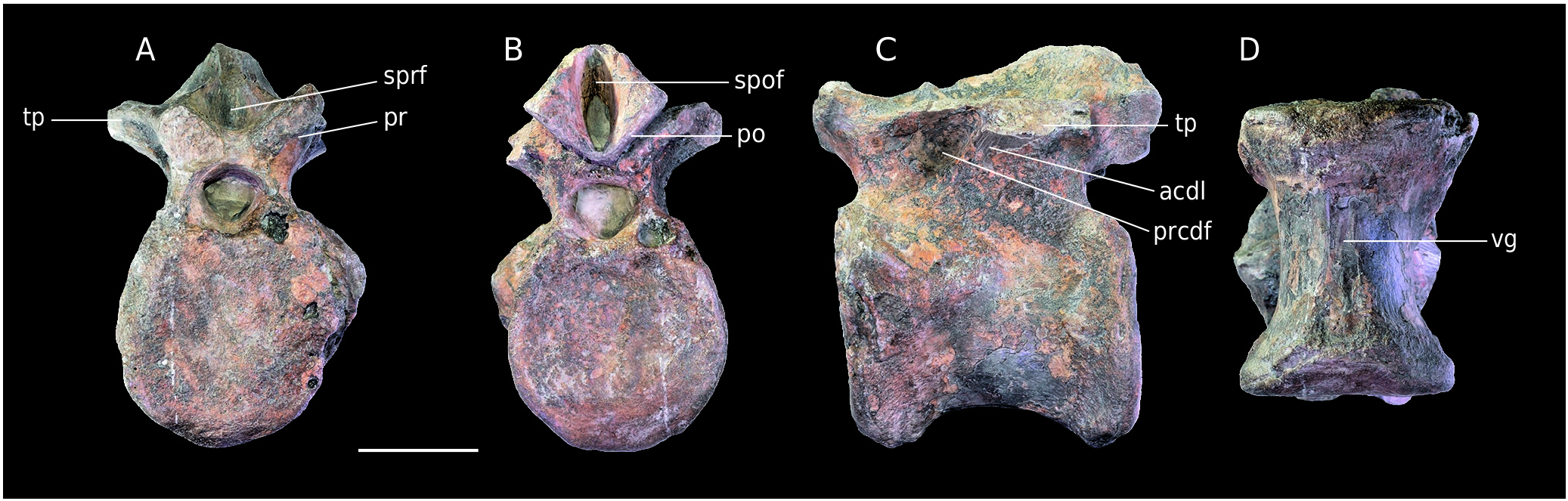
MPV 2020.1.7. Only the centrum of the vertebra is preserved ( Fig. 15A View FIG ). The left laterodorsal margin and the borders of the articular facets are eroded. The parapophyses are not visible. Thus, the preservation state prevents a clear orientation of the bone. The slightly greater depth of one side of the pleurocentral depression could indicate the front.
The centrum is platycoelous to slightly amphicoelous and is 110 mm long. It is much less concave laterally and ventrally than MPV 2020.1.7 and B5. This condition may be emphasised by erosion. Nevertheless, pleurocentral depressions are still much shallower. The right neurocentral suture is flat and clearly visible so that the individual may have been a juvenile. The neural canal is visible only medially. A 4 mm wide oval foramen pierces its centre.
The centrum shape seems to indicate a middle to posterior dorsal vertebra, but the absence of the parapophyses prevents a more precise positioning. Moreover, the state of preservation is too poor for a taxonomic assignment. And yet, this clade is use here more as a baseline, taking into account that the group is far more represented than others during Middle to Late Jurassic, especially in Europe ( Rauhut et al. 2016). MPV 2020.1.7 is different in shape with a transversally wider centrum bearing shallower lateral depression than MPV 2020.1.10 and B5 so that it may indicates a different taxon, tentatively referred to Tetanurae MPV V869. The vertebra was already described by Plasse & Buffetaut (2016) and defined as a thirteenth and last dorsal vertebra belonging to an allosauroid ( Fig. 15C, D View FIG ).
According to Madsen (1976), the last presacral vertebra of Allosaurus is the fourteenth.The number of cervical and dorsal vertebrae of the theropod have been discussed in Evers et al. (2015) so that it is assumed that Allosaurus had, as many other basal tetanuran rather 10 cervicals and 13 dorsals instead of 9 and 14. Then, Plasse & Buffetaut (2016) made a positioning correction with the Allosaurus fragilis vertebra used as comparison and figured ( Plasse & Buffetaut 2016: Fig. 2 View FIG ) that represents the fourteenth of Madsen (1976) but numbered as the thirteenth which indicate the last presacral vertebra. Parapophyses are potentially visible in the shape of an eroded area lateral to the centroprezygapophyseal fossae. If that is correct, they were located at the anterior base of the diapophyses and at the same level as the prezygapophyses, which supports the already established positioning.The specimen is therefore a last dorsal/ thirteenth vertebra.Despite the previous detailed description, the specimen was not compared with Megalosaurus bucklandii , particularly with a posterior dorsal vertebra of the paralectotype (OUMNH J.13577), which has important similarities with MPV V869. Among them is the flat area on the ventral side of the centrum, also present in Torvosaurus ( Britt 1991) . This feature is also present in Streptospondylus ( Allain 2001). The quite deep pleurocentral depression on each side of the vertebra is similar to Megalosaurus as well as to other megalosauroids such as Torvosaurus (e.g. Britt 1991). In the posterior dorsals of Allosaurus and Sinraptor , this structure is absent or less deep ( Madsen 1976; Currie & Zhao 1994). Regarding the neural arch, the “ridge” present in Megalosaurus bucklandi (OUMNH J.13577), which splits the “infrapostzygapophyseal” fossa ( Benson 2010), is also present and very similarly placed in MPV V869. This structure is the centropostzygapophyseal lamina anteriorly framing the centropostzygapophyseal fossa ( Wilson 1999; Wilson et al. 2011).
On the basis of these new observations, the assignment to Allosauroidea is less certain. The vertebra also bears similarities with Megalosaurus bucklandi and other megalosauroids. Thus, MPV V869 may also belong to the latter group. As a result, the specimen is referred to an indeterminate Tetanurae .
Caudal vertebrae
Material. Anterior caudal vertebrae NMH2, anterior to middle caudal vertebra B7 ( Fig. 16 View FIG ). Successive middle caudal vertebrae MPV 2020.1.4, MPV 2020.1.8 and MPV 2020.1.5 ( Fig. 17 View FIG ). Middle to posterior caudal vertebrae APVSM2 ( Fig. 18 View FIG ), MPV V795 and MPV 2020.1.6 ( Fig. 17 View FIG ); terminal caudal vertebra MPV 2020.1.12.
Stratigraphy. Marnes de Dives, Callovian marls according to the Nicolet collection catalogue made by Max Bülow and Michel Rioult (NMH2). Oxfordian or Callovian marls (B7, APVSM2, MPV 2020.1.4, MPV 2020.1.5, MPV 2020.1.6, MPV 2020.1.8 and MPV V795). Oolithe Ferrugineuse de Villers according to the ferruginous ooliths of the matrix. Lower Oxfordian ( MPV 2020.1.12).
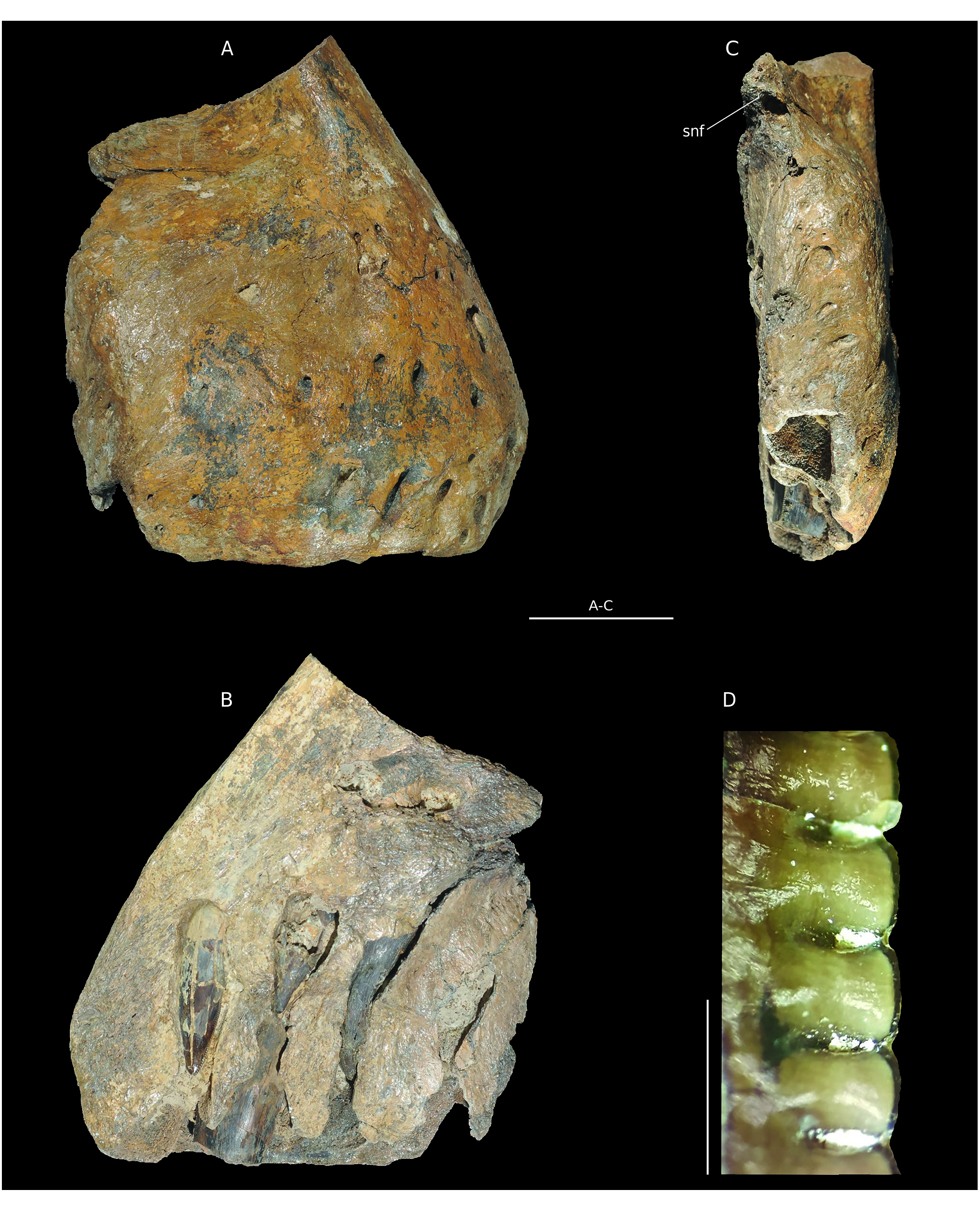
NMH2. This vertebra from the Nicolet collection was already described by Buffetaut in 1994. It was interpreted as an anterior caudal vertebra belonging to an indeterminate theropod. One of the remarkable features of the specimen is the “triangular depression” posterior to the prezygapophyses ( Buffetaut 1994b). This structure is laterally framed by a thick anterior centrodiapophyseal lamina and dorsally by the prezygodiapophyseal lamina and could be identified as the prezygapophyseal centrodiapophyseal fossa ( Wilson et al. 2011). Its position, like that of the transverse process; is quite posterior. The anterior centrodiapophyseal lamina is reduced and the fossa absent in Eustreptospondylus ( Sadleir et al. 2008) . These structures are not visible in Allosaurus but very similar to the condition in Metriacanthosaurus parkeri (OUM.J.12144/11).
B7. The vertebra is quite well preserved ( Fig.16 View FIG ). The transverse process and the right prezygapophysis are broken, but their bases are still visible, as is the neural spine. The left anterior part of the centrum seems inflated and is shattered so that it forms a protuberance ( Fig. 16A View FIG ). The centrum is 105 mm long and 75 mm wide. Following Madsen (1976), these proportions match the caudal vertebra of Allosaurus fragilis anterior to the thirtieth. Indeed, posterior to this position the centra become significantly more elongated so that they become twice as long as wide. Moreover, the neural arch is quite high and not flattened, supporting an anterior positioning of the specimen. The centrum is still longer than high ( Fig. 16C View FIG ) in contrast with the anterior caudal vertebrae (until about the seventh). Although incomplete, the neural spine seems quite thick Therefore, B7 should be located between the seventh and sixteenth vertebrae, more posteriorly than NMH2. The centrum is platycoelous and ventrally concave. The ventral face bears a groove limited laterally by two thin ridges of bone, as well as chevron facets anteriorly and posteriorly ( Fig. 16D View FIG ). The neural canal is subtriangular, laterally compressed in its ventral part by an enlargement of the pedicels of the neural arch. This is similar to Dubreuillosaurus and Eustreptospondylus ( Sadleir et al. 2008). A thick anterior centrodiapophyseal lamina as well as a fossa reminiscent of NMH2, are visible. The transverse processes seem quite posteriorly and horizontally oriented. The left prezygapophysis bears a mediodorsally inclined facet. The spinoprezygapophyseal laminae are well defined and nearly horizontal. They are posteriorly oriented and merge with the base of the neural spine, forming a deep horizontal spinoprezygapophyseal fossa. The postzygapophyses are well preserved and bear lateroventrallymediolaterally inclined subrectangular facets ( Fig.16B View FIG ). The postzygapophyses frame the spinopostzygapophyseal fossa anteriorly. The latter is vertical, in contrast with the spinoprezygapophyseal fossa. The neural spine, of which only the base is preserved, is quite posteriorly located on the neural arch.
Both specimens bear similar features such as the “triangular depression” already noticed by Buffetaut (1994b). Therefore, these two vertebrae should belong to closely allied taxa. It is also possible that they belong to the same individual. A precise positioning of these specimens is not possible considering the number of caudal vertebrae of a theropod. Allosaurus fragilis , like many other theropods such as basal Tetanurae , has about fifty ( Allain & Chure 2002). Regarding the significant morphological variation within a theropod tail ( Madsen 1976), a taxonomic assignment for an isolated specimen cannot be precise beyond Tetanurae .
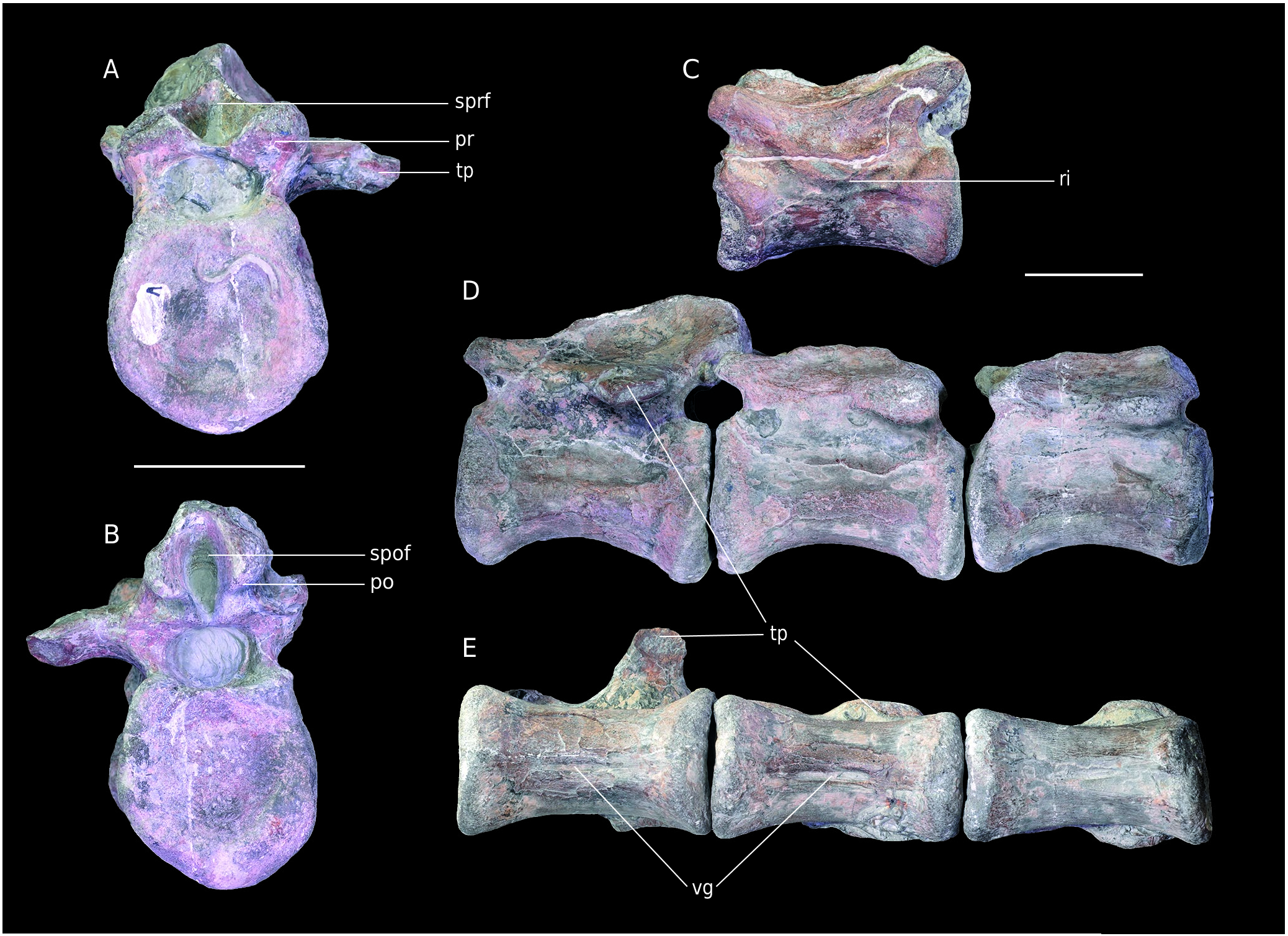
MPV 2020.1.4, 2020.1.8, 2020.1.5. These three vertebrae are anatomically successive and thus they are interpreted as belonging to the same individual ( Fig. 17D, E View FIG ). However, they were not found at the same time and place at the bottom of the cliffs by the Pennetier family. The matrix, the shape and the size as well as apposition of postzygapophyses to prezygapophyses and articular facets clearly indicate their connection. Their right side is covered with serpulids. MPV 2020.1.4 is the better preserved, its neural arch being clearly observable ( Fig. 17A, B View FIG ). Only the base of the right transverse process, the neural spine and right prezygapophysis remain. The neural arches of MPV 2020.1.8 and MPV 2020.1.5 are quite eroded. Both have quite well preserved prezygapophyses but not postzygapophyses, the common base of which is not visible. These 3 vertebrae have slightly amphicoelous centra which are almost twice longer than wide. The size of the centrum gradually decreases slightly along the series. Their ventral face bears a 20 mm to 30 mm longitudinal groove ( Fig. 17E View FIG ). The neural arch is flattened as compared to B7, but still well developed. As in Poekilopleuron bucklandi Eudes-Deslongchamp, 1837 , the transverse processes are in the posterior half of the vertebrae ( Allain & Chure 2002). The spinoprezygapophyseal and spinopostzygapophyseal fossae (the latter is only visible on MPV 2020.1.4) have a similar shape as in B7 which suggests a posterior development of the neural spine. The presence of transverse processes indicates that these vertebrae are anterior to the “transition point” as defined by Russell (1972). These structures are absent posterior to this point. In Allosaurus fragilis , it corresponds to the twenty-seventh vertebra ( Madsen 1976; Allain & Chure 2002). Based on the quite elongated centra as well as the rather low neural arches, these bones should be placed between the sixteenth and twenty-seventh vertebrae. The quite high position of the diapophyses could restrict the interval to the first twenty vertebrae. However, this positioning is questionable because of the isolated nature of the bones as well as the morphological variation within theropods.
APVSM2. the vertebra is quite eroded, especially on its right lateral side. Thus, most of the preserved structures are worn ( Fig. 18 View FIG ). The bases of the pre- and postzygapophyses as well as the left transverse process are visible ( Fig. 18A, B View FIG ). Only an abraded ridge -different from a natural bone surface- allows to discern the placement of the neural spine. The specimen has the same dimensions as MPV 2020.1.8 but is larger than MPV 2020.1.5, which indicates a different individual. The centrum is highly concave posteriorly and does not bear a ventral groove ( Fig. 18D View FIG ), as in Eustreptospondylus ( Sadleir et al. 2008) . This structure is, however, present in many theropods on their caudal vertebrae ( Madsen 1976). The neural arch is also different from that of the previous specimens. What remains of the base of the transverse process indicates a low position, below the neural canal ( Fig. 18C View FIG ). This is similar to caudal vertebrae just before the “transition point” (22 to 26 in Allosaurus fragilis Allain & Chure 2002 ). In contrast with the posterior position of the neural spine in MPV 2020.1.4, 8, 5, here it seems to expand all along the vertebra.
MPV V795 and MPV 2020.1.6. these vertebrae are quite damaged and cracked on the neural arch. Thus, a detailed description is not possible, and they are not figured. MPV 2020.1.6 is the better preserved. Like APVSM2, there is no ventral groove on the centrum. They present ridges or crests more or less notable instead of true transverse processes ( Fig. 17C View FIG ). These are eroded but allow to position the vertebrae after the “transition point”. However, MPV 2020.1.6 is larger than MPV 2020.1.5 and APVSM2, and thus it indicates a third individual.
Finally, the analysis of all these specimens suggests that there are at least 3 individuals. The variable morphology of vertebrae along a caudal series does not allow to determine whether they belong to different taxa. However, some specific features on some of the vertebrae, such as the ventral groove of MPV 2020.1.4, 8, 5, support that hypothesis. At least, MPV 2020.1.12 from the Oolithe Ferrugineuse de Villers, the stratigraphic origin of which is different from that of the other caudal vertebrae, indicates a fourth individual.

| MPV |
Museo Paleontologico Municipal de Valencia |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.