
Parastacus fluviatilis Ribeiro & Buckup
|
publication ID |
https://doi.org/10.11646/zootaxa.4158.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:9B375922-0D2B-4D92-99A1-14CAA259FBAB |
|
DOI |
https://doi.org/10.5281/zenodo.5691692 |
|
persistent identifier |
https://treatment.plazi.org/id/03EC87DA-DB3C-F342-FF4D-F97AFB1AFB00 |
|
treatment provided by |
Plazi |
|
scientific name |
Parastacus fluviatilis Ribeiro & Buckup |
| status |
sp. nov. |
Parastacus fluviatilis Ribeiro & Buckup View in CoL sp. nov.
( Figs. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Zoobank: urn:lsid:zoobank.org:act:7DB07615-B8C7-466A-9E2D-6C4DABCB233E
Holotype. Ƌ, Brazil, Rio Grande do Sul, São José dos Ausentes, Apuaê-Inhandava Basin, Silveira river ( 28°35’54.45”S; 49°59’1.36”W), 07/IX/1999, col. L. & T. Buckup ( MZUSP 34288 View Materials ). GoogleMaps
Paratypes. 1–3: Brazil, Rio Grande do Sul—two Ƌ and one ♀, same data as holotype ( UFRGS 2704 View Materials ) GoogleMaps ; 4–5: two Ƌ, São José dos Ausentes, affluent of Silveira river , Fazenda Potreirinhos ( 28°35’53.40”S; 49°58’55.99”W), 28/IV/1996, col. P.B. Araujo ( UFRGS 2294 View Materials ) GoogleMaps ; 6–7: two Ƌ, São José dos Ausentes, Fazenda Potreirinhos , 02/V/ 1997, col. O.R. Naches ( UFRGS 2295 View Materials ) ; 8–9: two Ƌ, São José dos Ausentes, affluent of Silveira river , Fazenda Potreirinhos ( 28°35’53.40”S; 49°58’55.99”W), 14/V/2014, col. K.M. Gomes, F.B. Ribeiro & D.C. Kenne ( UFRGS 6195 View Materials ) GoogleMaps ; 10–11: two Ƌ, São José dos Ausentes, margin of Silveira River , near the gap of Silveira and Divisa rivers, Fazenda Potreirinhos ( 28°36’21.29”S; 49°58’41.74”W) 15/V/2014, col. K.M. Gomes, F.B. Ribeiro & D.C. Kenne ( UFRGS 6107 View Materials ) GoogleMaps ; 12: one ♀, São José dos Ausentes, Marco river ( 28°36’42.99”S; 49°55’11”) ( UFRGS 2236 View Materials ) ; 13– 15: two Ƌ and one ♀, Bom Jesus, Fazenda Sr. Argemiro P. Borges, 28/I/1979, col. O. Camargo (UFRGS 1363).
Comparative material analyzed. Brazil, Rio Grande do Sul: Parastacus pilimanus —one Ƌ and one ♀, Rio Grande, Taim , 09/VI/1975, col. L. Buckup ( UFRGS 277 View Materials ) ; one ♀, Dom Pedrito , 05/III/1957, col. C.P. Coreto ( UFRGS 1374 View Materials ) ; one Ƌ and one ♀, São Gabriel ( 30°34’16.86’’S; 54°29’42.22’’W), 21/IX/2012, col. K.M. Gomes ( UFRGS 5756 View Materials ) GoogleMaps ; one Ƌ, Ibirapuitã river, Alegrete , 25/II/1982, col. B. Irgang ( UFRGS 542 View Materials ) ; Parastacus brasiliensis —three Ƌ, Mariana Pimentel ( 30°20’41”S; 51°33’55”W), 12/IV/2010, col. W. Beduchaud, K.M. Gomes & S. Santos ( UFRGS 4890 View Materials ) GoogleMaps ; one Ƌ and one ♀, Porto Alegre, Praça da Vila Jardim Renascença , Zona Sul ( 30°5’79”S; 51°11’30.62”W), 10/VII/2013, col. K.M. Gomes, F.B. Ribeiro & G.C. Dalló ( UFRGS 5860 View Materials ) GoogleMaps ; Santa Catarina: P. laevigatus —one ♀ and five juveniles, Estrada da Cidra, Chacara dos Ipês, Joinville , 05/08/1961, col. Rosenberg ( UFRGS 1369 View Materials ) .
Etymology. The Latin epithet fluviatilis alludes to the riverine habitats where this species was found. We suggest the common name “the highland streams crayfish” for this new species.
Diagnosis. Narrow front with short triangular rostrum. Rostral apex shaped as inverted “U”, with blunt spine. Postorbital ridges present and easily distinguished only in anterior portion. Cervical groove strongly V-shaped. Areola narrow and with no lateral elevation on branchiostegite grooves. Cutting-edge surface of fixed finger and dactylus with dense cover of simple and pappose setae. Telson subtriangular with small blunt lateral spines. Mandible with caudal molar process unicuspidate with one big cephalodistal cusp. Ventral surface of lateral process of thoracic sternites 6 and 7 strongly concave.
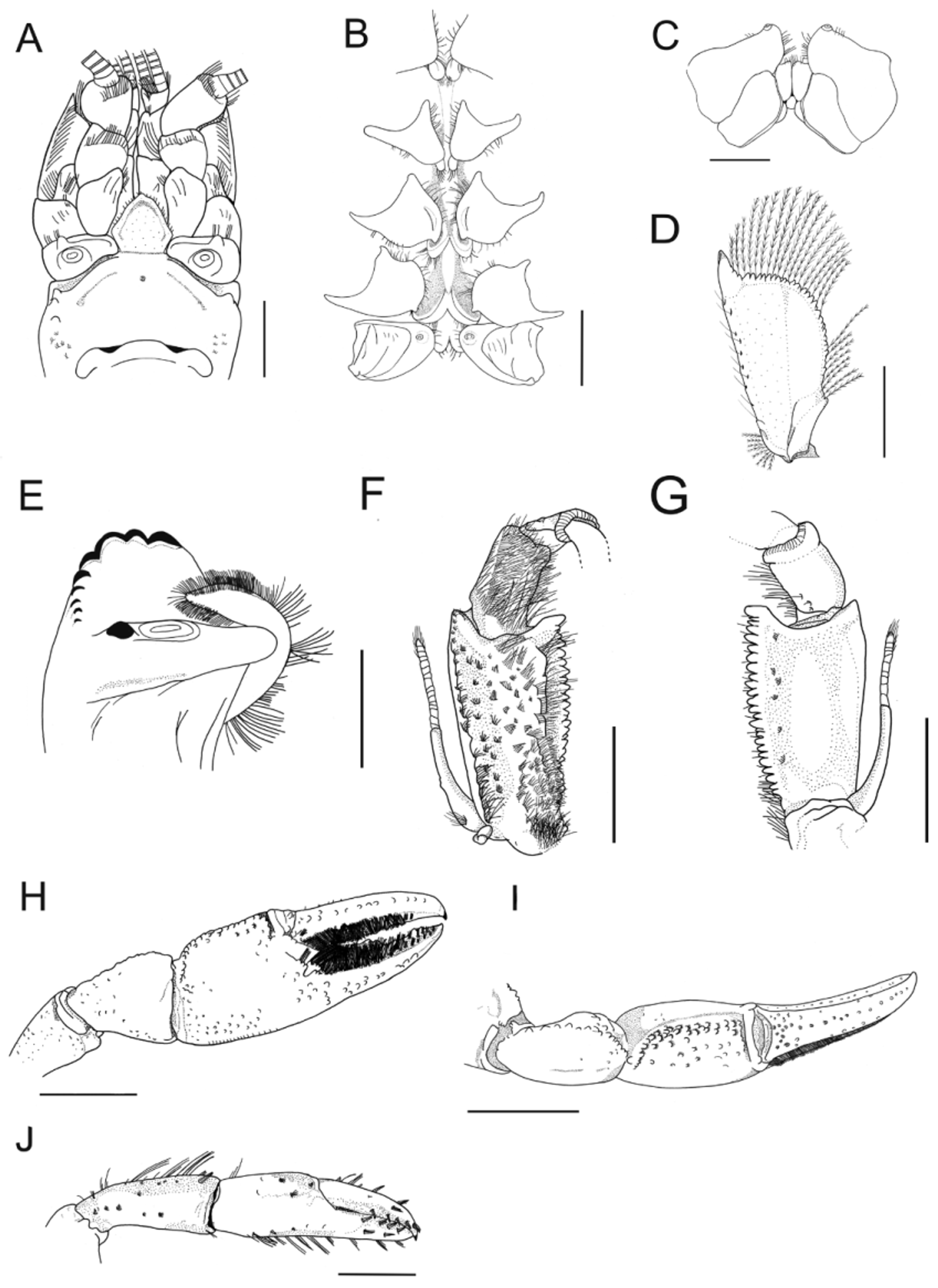
Description. Rostrum: triangular, wider than long (RL 98.7% of RW), short (12.4% of CL), reaching middle of second article of antennular peduncle ( Fig. 3 View FIGURE 3 A, B, C). Dorsum slightly concave, apex inverted “U”-shaped, ending in straight blunt spine. Few plumose setae on lateral margins. Rostral sides slightly convergent and rostral basis slightly divergent. Carinae almost straight, prominent and wide, extending back to carapace, surpassing rostral basis ( Fig. 3 View FIGURE 3 B, C).
Cephalon: Carapace lacking spines or tubercles. CeL 63.9% of CL. Eyes small (CMW 64.4% of OW); suborbital angle 90°, unarmed ( Fig. 3 View FIGURE 3 C). Front narrow (FW 40% of CW). Postorbital carinae longer than rostral carinae (RCL 62.6% of POCL), conspicuous anteriorly and barely discernible posteriorly. Lateral cephalic edge with sparse setation.
Thorax: carapace laterally compressed, deep and narrow (CD 50.8% of CL; CW 44.1% of CL). Cervical groove strongly V-shaped. Branchiocardiac grooves conspicuous and without carina ( Fig. 3 View FIGURE 3 A). Areola narrow, 3.1x as long as wide (29.3% of CL) ( Fig. 3 View FIGURE 3 A).
Abdomen: lacking spines or tubercles, long and wide (AL 79% of CL; AW 80.3% of CW), smooth, with few small setae on pleural margins ( Fig. 3 View FIGURE 3 A, D). Pleural somites with rounded posterior margins. S1 pleurae with small distal lobe not overlapped by S2 pleurae. S2 pleurae with shallow groove parallel to margin, high and moderately long ( Fig. 3 View FIGURE 3 E, F).
Tailfan: telson calcified in the proximal portion and weakly calcified in the distal margin, subtriangular, longer than wide (TeW 81.6% of TeL), with small blunt spines on lateral margins; rounded distal margin with abundant long plumose setae and short simple setae. Dorsal surface with tufts of short setae and a dorsomedian longitudinal sulcus ( Fig. 3 View FIGURE 3 G). Uropod protopod bilobed, with rounded and unarmed margins; proximal lobe largest. Exopod lateral margin unarmed, mid-dorsal carina weakly prominent, ending in small spine. Transverse suture (diaeresis) straight, with two dorsolateral spines (outer) and one dorsolateral spine (inner) on each of right and left exopodites. Endopod, mid-dorsal carina weakly prominent, ending in spine, lateral margin with small spine at level of exopod transverse suture ( Fig. 3 View FIGURE 3 G).
Epistome: anterolateral section with three marginal tubercles, posteriormost tubercle smallest; also with single keel and small circular median concavity. Posterolateral section with cluster of squamose setiferous tubercles. Anteromedian lobe irregularly pentagonal, 1.03x longer than wide, reaching midlength of antepenultimate article of antennal peduncle; lateral margins keeled; ventral surface slightly concave and basis deeply grooved ( Fig. 4 View FIGURE 4 A).
Thoracic sternites: SLP4 small and very close to each other, median keel present and not inflated; SLP5 smallest and close to each other, median keel present and not inflated; SLP6 larger than SLP4 and SLP5 and with concave surface, median keel inflated; SLP7 largest and with surface strongly concave, median keel inflated and rhomboidal, bullar lobes absent; SLP8 smaller than SLP6 and concave, median keel absent, vertical arms of paired sternopleural bridges widely separated, bullar lobes separated and clearly visible ( Fig. 4 View FIGURE 4 B, C).
Antennule: internal ventral border of basal article with sharp spine ( Fig. 4 View FIGURE 4 A).
Antenna: when extended back reaching second somite of pleon. Antennal scale widest at midlength, reaching midlength of third antennal article, ASW 40.8% of ASL ( Fig. 4 View FIGURE 4 A, D), lateral margin slightly curved, spine strong and distal margin emarginate. Coxa with weakly prominent carina above nephropore, with strong blunt dorsolateral spine. Basis unarmed ( Fig. 4 View FIGURE 4 A).
Mandible: cephalic molar process molariform, caudal molar process unicuspidate with one big cephalodistal cusp. Incisive lobe with nine teeth. The third tooth from the anterior is the largest. ( Fig. 4 View FIGURE 4 E).
Third maxilliped: ischium bearing several setiferous puctuations but with numerous short smooth simple setae on outer margin and on ventrolateral surface, but some tufts presents longer setae ( Fig. 4 View FIGURE 4 F) and the dorsal surface presents few setiferous punctuations ( Fig. 4 View FIGURE 4 G). Merum ventral surface entirely covered by long smooth simple setae ( Fig. 4 View FIGURE 4 F). Crista dentata of right and left ischium each with 28 teeth. Merus, entire ventral surface densely covered with simple setae. Exopod longer than ischium, with flagellum reaching proximal margin of merus ( Fig. 4 View FIGURE 4 F, G).
First pair of pereiopods (chelipeds): large and subequal, laterally flattened (RPrT 23.2% of RPrL; LPrT 22.6% of LPrL) ( Fig. 3 View FIGURE 3 A). Ischium ventral surface with 11 tubercles. Merus: right merus (RML) 48.2% of propodus length (RPrL); left merus (LML) 49.5% of propodus length (LPrL); ventral surface with two longitudinal series of tubercles: inner series with 15 tubercles, outer 12 and mesial 16, arranged irregularly on right merus; inner series bearing 11 tubercles, outer 13 and mesial 15, arranged irregularly on left merus. Dorsal and midventral spines absent. Carpus with dorsomedial surface divided longitudinally by shallow groove ( Fig. 3 View FIGURE 3 A; Fig. 4 View FIGURE 4 I). Internal dorsolateral margin with row of tubercles, increasing in size distally; inner surface with 1–2 small mesial tubercles. Carpal spine absent ( Fig. 4 View FIGURE 4 I). Propodus width (RPrW and LPrW) 39.61% of length in right cheliped and 41.75% in left cheliped. Dorsal surface of palm with two rows of tubercles ( Fig. 4 View FIGURE 4 H, I). Inner margin bearing few small tubercles. Ventral surface bearing two rows of squamose tubercles, reaching beginning of fixed finger ( Fig. 4 View FIGURE 4 H). Dactylus: moving subvertically, right dactylus (RDL) 54.6% of propodus length (RPrL), left dactylus (LDL) 58.4% of left propodus (LPrL); dorsal surface without tubercles, but with rows of bristle tufts ( Fig. 4 View FIGURE 4 I). Cutting edge of fingers covered with tufts of pappose setae; fixed finger with eight teeth and dactylus with nine teeth, uniformly distributed, decreasing in size distally ( Fig. 4 View FIGURE 4 H, I).
Second pair of pereiopods: ventral and dorsal surface of carpus, propodus and dactylus with sparse covering of simple setae ( Fig. 4 View FIGURE 4 J).
Gonopores: Presence of both genital apertures on coxae of third and fifth pairs of pereiopods. Female gonopores semi-ellipsoidal (maximum diameter 1.5 mm) with well-calcified membrane. Male gonopores rounded, opening onto apical end of a small, fixed, calcified and truncated phallic papilla, close to inner border of ventral surface of coxae of fifth pair of pereiopods. Male cuticle partition present ( Fig. 4 View FIGURE 4 B).
Branchial count: 20 + epr + r. Branchial arrangement follows the same described by Huxley (1879) and Hobbs (1991) with the epipodite of the first maxiliped with rudimentary podobranchia filaments.
Measurements. Holotype male, CL 38.66 mm and TL 76.66 mm. In type series, CL ranging from 18.25 to 40.88 mm (27.96 ± 7.04 mm). Female paratypes larger than males. FW/CW: 0.44 ± 0.04 (min: 0.39; max: 0.51). RL/RW: 1.00 ± 0.19 (min: 0.74; max: 1.35). CMW/OW: 0.67 ± 0.11 (min: 0.5; max: 0.83). Postorbital carina longer than rostral carina in all specimens analyzed. CW/AW: 1.16 ± 0.12 (min: 1.04; max: 1.51). AuW/RW: 1.18 ± 0.35 (min: 0.71; max: 1.72).
Color of live specimens. Rostrum greenish brown. Cephalothorax anterior region brown, lateral region light brown. First pair of pereiopods brown with greenish-brown fingers and whitish setae coverture on cutting edge. Pereiopod pairs 2–5 light brown. Dorsal pleon brown to greenish with light-brown to greenish-brown marks on lateral margins. Tailfan brown with shades of light brown to greenish brown on dorsal surface of telson ( Fig. 5 View FIGURE 5 E, F).
Remarks. Female paratypes differ from holotype in their larger sizes, narrower rostrum, larger pleurae of abdominal somites ( Fig. 3 View FIGURE 3 D, E, F) and shorter chelipeds. Allometric differences between males and females, as found in other crayfishes, are possible ( Reynolds 2002), but were not analyzed here. All paratypes presents both masculine and feminine gonopores in the same individual. Male paratypes also present female gonopores semiellipsoidal (average maximum diameter 1.24 ± 0.37 mm) covered by a calcified membrane. Female paratypes presents female gonopores ellipsoidal (average maximum diameter 1.88 ± 0.31 mm) covered by a thin and less calcified membrane. Male gonopores are very similar in males and females.
Parastacus fluviatilis View in CoL sp. nov. is morphologically similar to P. pilimanus (von Martens, 1869) View in CoL and P. laevigatus View in CoL in having the cutting edge of the cheliped fingers covered with long setae ( Buckup & Rossi, 1980). It differs from P. pilimanus View in CoL in that the latter has a longer rostrum, rows of verrucous tubercles on the dorsal surface of the dactylus of the first pereiopods/chelipeds ( vs. dorsal surface without tubercles, but with rows of bristle tufts in P. fluviatilis View in CoL sp. nov.), and two sharp spines on the right side of the coxa of the antennal peduncle above the nephropore (please, follow the example of previous comparison). And P. fluviatilis View in CoL differs from P. laevigatus View in CoL mainly in the absence of a carpal spine in chelipeds, the post orbital ridges well prominent, cervical groove strongly V-shaped and larger body sizes. Parastacus fluviatilis View in CoL sp. nov. differs from all other Parastacus View in CoL species in having a subtriangular telson, postorbital ridges well distinguished only in anterior portion and a strongly concave ventral surface of the lateral process of thoracic sternites 6 and 7.
Habitat and ecology. Parastacus fluviatilis View in CoL sp. nov. was collected at altitudes above 1,300 m in streams of the Serra Geral plateau. This physiographic region is called Campos de Cima da Serra ( Fig. 5 View FIGURE 5 ), i.e., the Brazilian Subtropical Highland Grasslands ( Iganci et al. 2011). The streams and rivers in this region characteristically have stone and pebble bottoms. They are bordered by few riparian forests, composed mainly of undergrowth and shrubby vegetation ( Bond-Buckup 2008).
The specimens were collected with traps in first- and third-order streams, and in backwaters, with dip nets. Shallow burrows were found on the streambanks, usually with one simple opening; when present, the chimney was short ( 4–5 cm high) ( Fig. 5 View FIGURE 5 C). P. fluviatilis sp. nov. is ecologically similar to P. brasiliensis in that both inhabit streams and build shallow burrows in the streambanks ( Buckup & Rossi 1980).
Distribution. Parastacus fluviatilis sp. nov. appears to have an extremely limited distribution, being found only in mountain streams of northeastern Rio Grande do Sul, in São José dos Ausentes and Bom Jesus municipalities ( Fig. 2 View FIGURE 2 ). The main streams where the species occurs are Silveira and Marco, tributaries in the Apuaê- Inhandava hydrographic basin, Uruguay Hydrographic Region ( Justus 1990).
Conservation status. The EOO was estimated at approximately 1,140 km ² based on the Otto Bacia shape level 6 ( ANA 2006), indicating that this species can be included in the Endangered—EN category, in which the EOO is less than 5,000 km ² (IUCN). The species is categorized as EN under subitem "a": it is known to occur at no more than 5 locations, as established from its presence in the sub-basins (Silveira and Marco rivers), and local threats; and subitem "b" (iii): continuing decline in quality of habitat by the threats observed in the field, such as deforestation of riparian vegetation for cattle ranching, planting of exotic species (e.g., Pinus sp.), agriculture (e.g., sweet potato, Ipomoea ) and the presence of the exotic rainbow trout Oncorhynchus mykiss Walbaum, 1972 . We suggest that the conservation status of this species be classified as ENDANGERED B1 ab(iii).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Parastacus fluviatilis Ribeiro & Buckup
| Ribeiro, Felipe Bezerra, Buckup, Ludwig, Gomes, Kelly Martinez & Araujo, Paula Beatriz 2016 |
P. pilimanus
| von Martens 1869 |