
Peignecyon felinoides, Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella, 2019
|
publication ID |
https://doi.org/ 10.2478/if-2019-0025 |
|
persistent identifier |
https://treatment.plazi.org/id/03E7E248-FFF9-FF9F-846A-BCDAFB0FFAFC |
|
treatment provided by |
Diego |
|
scientific name |
Peignecyon felinoides |
| status |
sp. nov. |
Peignecyon felinoides n. sp.
Text-figs 1 View Text-fig , 2 View Text-fig , Pls 1–3
2003 Tomocyon nov. sp.; Fejfar et al., p. 167.
2016 Tomocyon (? nov. gen.), nov. sp.; Fejfar and Heizmann, p. 325.
H o l o t y p e. NM-Pv 11600 right mandible with p4–m1, alveoli for m2–m3 (Pl. 1), deposited in NMP.
P a r a t y p e. TU 73916 right M1 ( Text-fig. 2b View Text-fig ).
E t y m o l o g y. “felinoides” from feline, denomination used by Hürzeler (1940) for this group of Amphicyonidae .
Ty p e l o c a l i t y. Tuchořice, the Czech Republic.
A g e. Early Miocene (MN 3).
D i a g n o s i s. The same as for the genus.
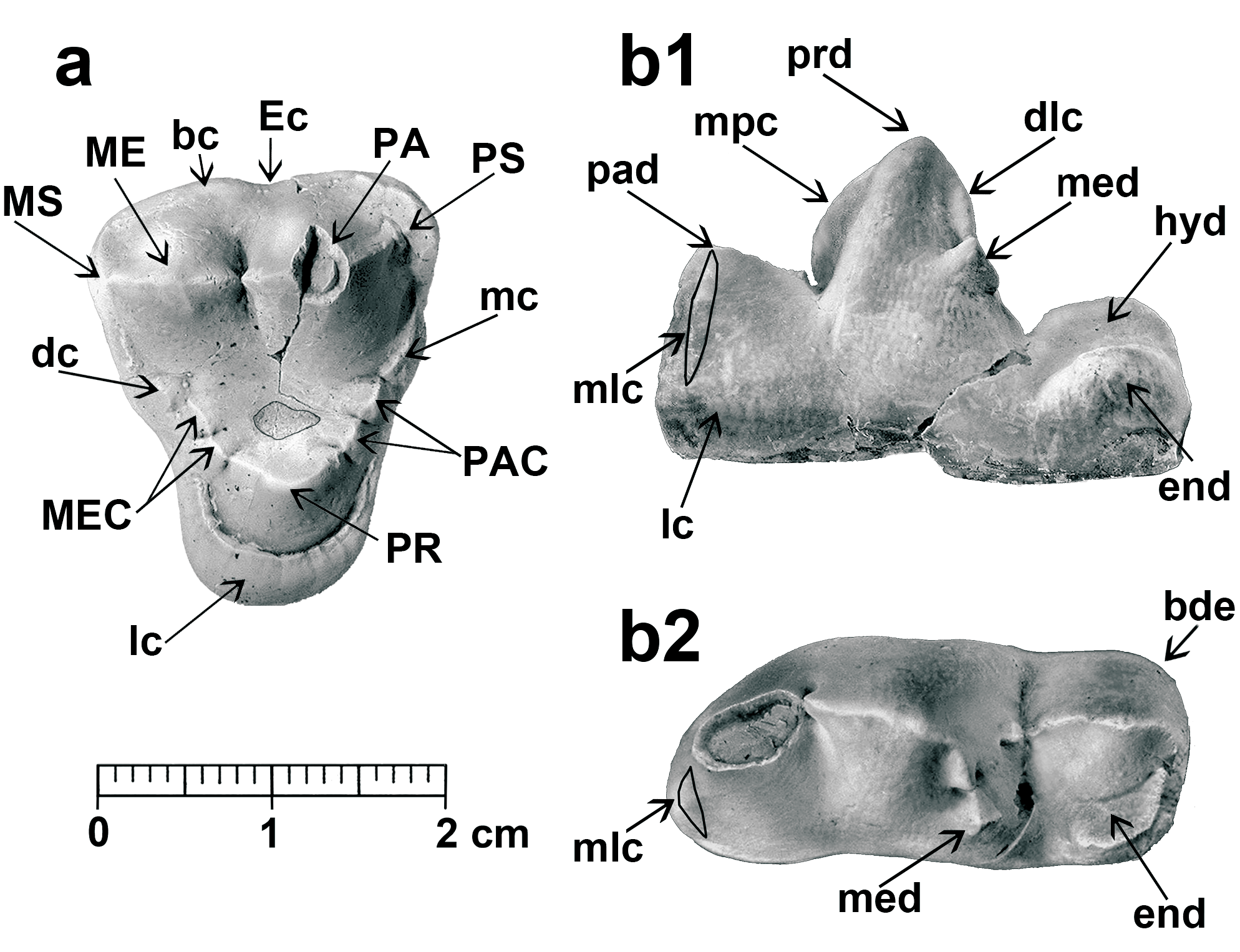
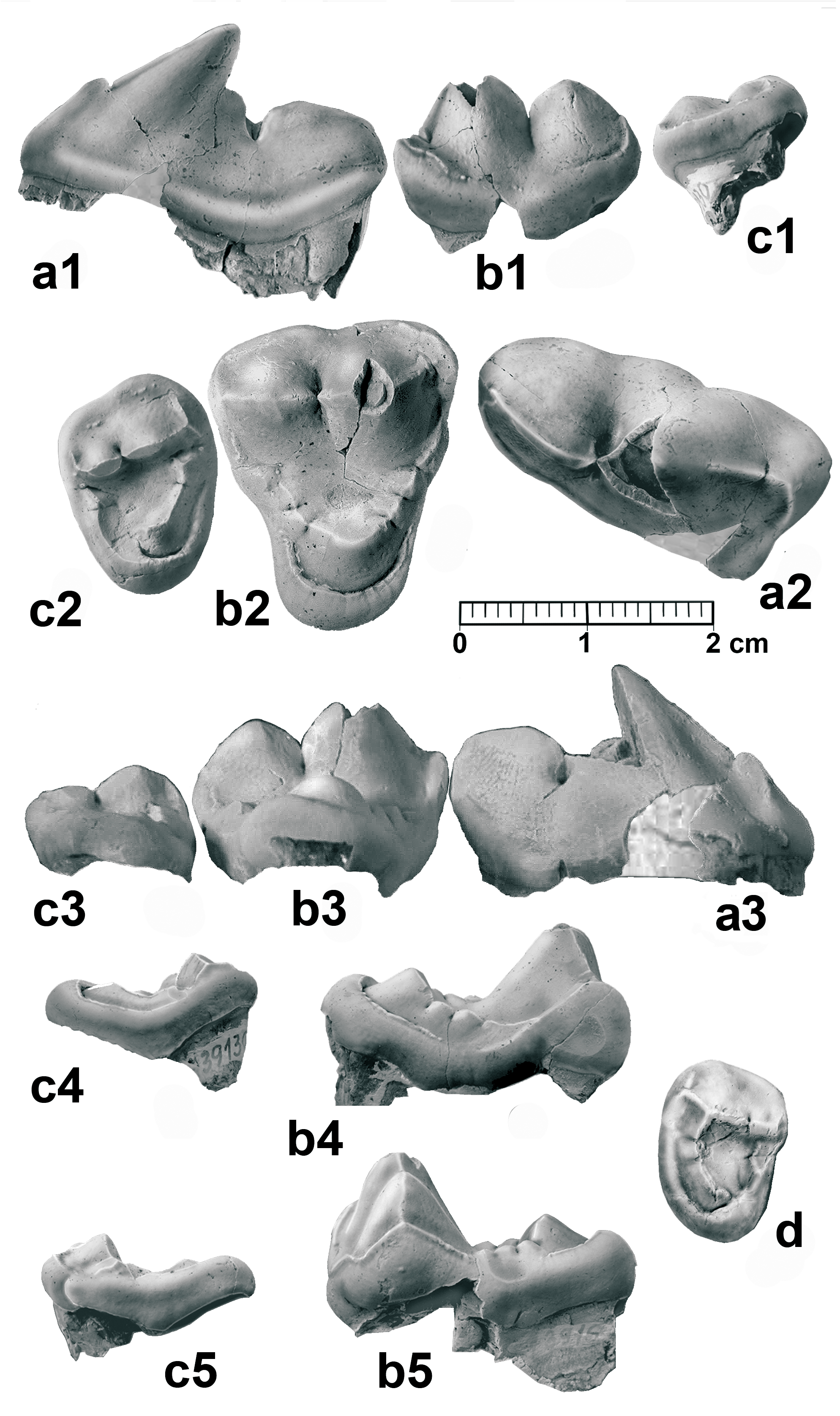
D e s c r i p t i o n. Mandible and lower dentition. NM- Pv 11600 right mandible with c, p1 alveolus, p4–m1, and alveoli for m2–m3 (holotype, Pl. 1), the mandible is included in the sediment with the lingual wall exposed. The mandible ramus was partially broken, but the fragments remained close to the corpus mandibulae, and this part has thus been restored in the image (Pl. 1, Fig. 1a). The overall morphology is close to that of the wolf ( Canis lupus ), but is somehow shorter and more robust. There is a long diastema between the p4 and the alveolus for the p1 without the alveoli for p2–p3. The margo interalveolaris is sharp, and in lingual view it presents a concave profile (Pl. 1, Fig. 1a, b), common in species with premolar reduction and presence of a long diastema, as occurs in Thaumastocyon bourgeoisi (see Stehlin and Helbing 1925) or Pseudocyon ? astrei KUSS, 1962. The canine associated with the mandible is well developed, and presents a sharp and irregularly crenulated distal cristid (Pl. 1, Fig. 2). The p4 is small in relation to the m1, presenting a smooth paraconid. The protoconid is distally inclined, high and with a smooth mesial cristid. The distal cuspid is well individualized and the talonid is small. The lingual cingulum is somewhat more developed in the distal part of the tooth, where it is slightly expanded. The m1 is a sectorial tooth with moderately laterally compressed paraconid and protoconid. The paraconid mesial cristid is not totally vertical. The m1 metaconid is a small bulge in the middle of the distolingual protoconid cristid. The talonid is very short, and is mostly occupied by a tall and long hypoconid. The mesial cristid of the hypoconid is aligned with those of the protoconid and paraconid, forming the carnassial blade. The entoconid is relatively large and located parallel to the hypoconid, and both cuspids are separated by a very narrow valley. A small cristid in the hypoconulid position connects the entoconid with the hypoconid. The m2 is missing, but the alveoli are seen to present a moderate size. The alveolus for the m3 is very small, with one single root.
TU 73917 left mandible with p4–m2 and alveoli for p3 and m3 (Pl. 2, Fig. 1). The presence of a possible p3 is marked by a one-rooted alveolus, separated from the p4 by a small diastema. Both the p4 and the m1 are morphologically close to the holotype. The m1 metaconid is reduced to a small notch in the distolingual cristid of the protoconid. The m1 talonid is also very short, with a small entoconid situated in its distolingual corner. The distal end of the talonid is rectilinear. The cingula are very smooth. There is a deep vertical occlusion facet in the labial wall, between the protoconid and the hypoconid, as well as vertical wear facets on the labial wall of both the paraconid and the protoconid. The m2 exhibits a quadrangular shape, with a complete trigonid, although the paraconid is low and very reduced. Its labial cristid connects the mesial cristid of the protoconid, and appears to be labially attached to the metaconid, in such a way that a well-defined mesiolingual valley is formed among them. The protoconid is quite large, with a pyramidal morphology; it has three cristids, one mesial, another distal (parallel to the molar axis) and the third one lingual, contacting the metaconid. Its labial wall is basally widened. The metaconid is smaller and lower than the protoconid, and is slightly displaced distally. The talonid is short, with a low and wide hypoconid. The entoconid is also low, and is located in the distolingual corner. Between these two cusps, a narrow and relatively deep valley can be observed. The cingula are smooth. The presence of a small m3 is attested by a small distal alveolus, partially broken. The m3 is represented in some specimens TU 739118 (Pl. 3, Fig. 1) , TU 739111 (Pl. 3, Fig. 4); it is morphologically close to the m2, but with less defined cuspids and an overall smaller size .
Some morphological variations can be recognized in other specimens; for example, TU 7391147, a right mandible (Pl. 3, Fig. 3), lacks the parastylid in the p4 and its distal cusp is somewhat stronger; the m1 retains a small metaconid, and the mesial cristid of the paraconid is slightly displaced lingually and highly marked. Furthermore, the labial wall of the hypoconid is somewhat basally expanded, developing an incipient cingulum. The m2 represents quite a different morphotype, in particular due to the greater reduction of the talonid, which is almost exclusively built by the hypoconid; this morphology is also observed in the left m2 TU 939151 (Pl. 3, Fig. 5). The m1 of the specimen TU 739111 (Pl. 3, Fig. 4) has a small metaconid, but better developed than that of the previously mentioned specimens, as occurs with the mesial cristid of the paraconid, which is well-marked and tends to be in a more lingual position.
Upper dentition. TU 739130 right P4, elongated upper carnassial ( Text-fig. 2a View Text-fig ). The paracone and protocone are partially broken. There is a well-developed lingual cingulum. The parastyle is present and individualized, but still moderate in size. The protocone appears to be reduced, and its position is somewhat distal. The paracone is coniform, with a very marked mesial crista presenting a clear distal inclination. The metastyle is quite robust.
TU 73916 right M1 ( Text-fig. 2b View Text-fig ). The occlusal outline is an isosceles triangle with the labial wall rectilinear, with a smooth ectoflexus. The parastyle is quite well developed. The paracone is rather larger and higher than the metacone. The metastyle is small. The labial cingulum is high and strong at the base of both the paracone and metacone, but weaker in the ectoflexus area. The paraconule and the metaconule are subdivided and well developed. The paraconule comes into contact with a high cingulum, developed from the parastyle, whereas the metaconule connects with a small cusp associated with the metastyle cingulum, which is very weak lingually. The protocone is situated in a central position relative to the labial wall; it is strong, with a duniform shape and completely encircled by a strong labial cingulum, which reaches the bases of the paraconule and metaconule. It is thickened at its distolingual corner. The lingual part of the molar is clearly reduced compared to the labial one. In the distal wall there is a strong inflexion at the position of the metaconule, while the mesial wall is more rectilinear. The central valley of the trigone is deep, revealing a circular wear pattern in the centre, resulting from the occlusion with the m1 hypoconid. An additional right M1, TU 739129, is poorly preserved; it is somewhat smaller but morphologically similar to the other molar.
TU 739130 right M2 ( Text-fig. 2c View Text-fig ). It presents an oval shape and small size in relation to the M1. Its basal cingulum surrounds the complete molar, thickening at the distolingual corner. Both the paracone and the metacone are low and somewhat lingually displaced. Both parastyle and metastyle are small. The protocone is poorly developed; it is located in a central position and joined by a long crista to the paraconule. The metaconule is included in the distal cingulum, and does not make contact with the protocone. There is a small cusp lingual to the metacone, associated with the metastyle. The trigone valley is broad and flat. Specimen TU 739136 is also an M2, which is similar in shape to TU 739130, but smaller in size ( Text-fig. 2d View Text-fig ).
D i s c u s s i o n. The hypercarnivorous character of Peignecyon felinoides is unquestionable; it is manifested in the strong development of the sharp mandibular diastema, associated with the reduction of the anterior premolars, p4 high, with a distally inclined mesial cristid, carnassials with sectorial morphology, characterized by the reduction of the m1 metaconid and the strong development of the hypoconid, and in the elongation of P4 with an incipient parastyle. The M1 presents a triangular occlusal shape and a very reduced lingual area. The second molars, m2 and M2, respectively, are smaller in relation to the m1 and M1. Some of these characteristics are also found in Ysengrinia gerandiana ( Viret 1929a, Ginsburg 1965) from Saint Gérand le Puy ( France), early Miocene (MN 2). However, in the latter species, these hypercarnivorous characters are less developed. Among other traits, it possesses a less reduced lower premolar row with shorter diastemas; the p4 has the principal cusp smaller and more vertical, with no distal inclination of the mesial cristid; the m1 is more robust and retains a strong metaconid. Additionally, the talonid is wider than the trigonid, and its hypoconid and entoconid are comparatively less developed. These differences between both amphicyonids are noteworthy, and suggest a clear taxonomic separation between them.
Peignecyon felinoides differs from other species included in the genus Ysengrinia , such as Ysengrinia tolosana ( NOULET, 1876) and Ysengrinia depereti ( MAYET, 1908) , in its smaller dentition, a more gracile m1 with a more reduced metaconid, and the m2, which has lower and smaller cuspids. The absence of upper dentition for these taxa severely limits the comparisons. However, the differences in relation to P. felinoides are more pronounced than those indicated for Y. gerandiana .
The species Ysengrinia americana ( WORTMAN, 1901) is much better represented; it was originally described as Amphicyon , and included by Hunt (2002) in this European genus. The relationship between European and American species has been exhaustively discussed by this author, who finds obvious affinities between Y. americana and Y. tolosana , and between the latter two and Y. gerandiana . However, there are notable differences in the upper dentition, which obscure these relationships. The M1 of Ysengrinia americana shows a strong reduction of the lingual area, similar to that observed in Y. gerandiana and P. felinoides , but the lingual cingulum does not completely encircle the protocone as it does in these two European species. On the contrary, this cingulum is restricted to the distolingual margin, so that the mesial wall of the protocone forms part of the wall itself, and its cingulum is only a small ridge. Additionally, the M1 of P. felinoides has buccal cusps that are much higher than in the American species, and the paraconule and metaconule are coniform and subdivided, whereas in Y. americana they form crests and are attached to the protocone. In the M2, the differences are even more pronounced; in Y. americana the molar is morphologically close to the M1, but the protocone is completely surrounded by the lingual cingulum, which is comparatively better developed. In Y. gerandiana and P. felinoides , the M2 is very reduced lingually, and the molar is therefore oval, and the protocone is very reduced. Thus, in both European species, the M2 has a similar overall shape of the M3 to Y. americana and other amphicyonid species. This fact can be related to the strong tendency of Y. gerandiana and P. felinoides to reduce the crushing molar, whilst in Y. americana , this feature is less pronounced. The lower dentition of P. felinoides also differs from that of Y. americana in the greater gracility of the p4 and the m1. Considering the current data, we conclude that Y. americana cannot be included within the genus Ysengrinia . Consequently, Y. tolosana and Y. depereti , only represented by their lower dentition, also remain in an uncertain systematic position, awaiting documentation on their upper dentition.
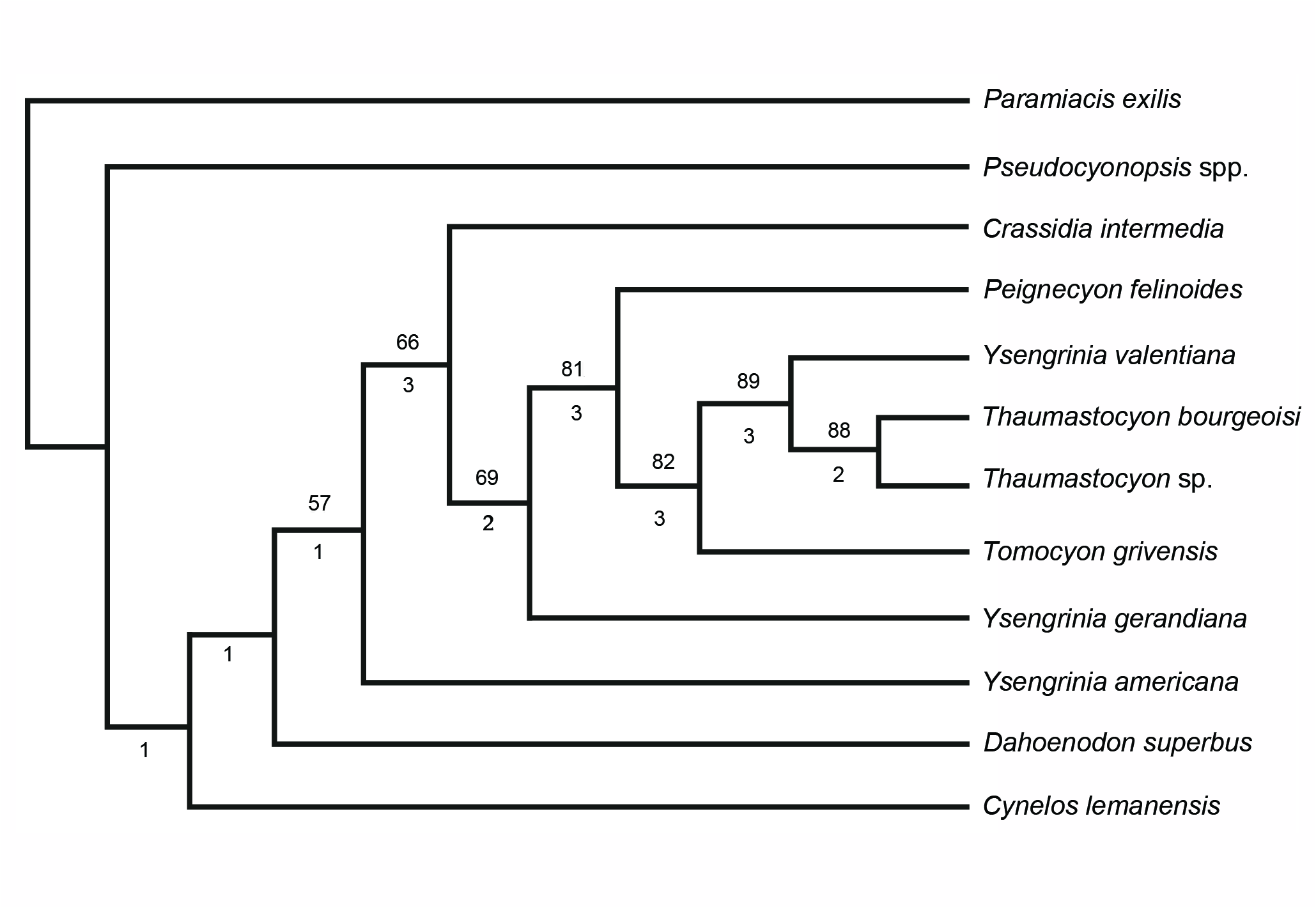
Although in our analysis, Y. americana could be considered as the sister group of the Thaumastocyoninae ( Text-fig. 3 View Text-fig ), alternatively, it could be related to other hypercarnivorous Amphicyoninae that have retained a well-developed M2 and a reduced p4 ( Peigné et al. 2008, Morales et al. 2016). These Amphicyoninae require an indepth taxonomic and systematic review, but this is not the objective of the present paper.
A second genus Crassidia was included close to Ysengrinia in the Ysengrini tribe by Heizmann and Kordikova (2000). Peignecyon felinoides differs from other species grouped in the genus Crassidia ( Heizmann and Kordikova 2000) in the general gracility of its dentition. Significant differences are also seen in the morphology of the M1, which in the holotype of Crassidia intermedia from Ulm has a very developed lingual area, as occurs in the maxilla fragment of Selles-sur-Cher ( Heizmann and Kordikova 2000: pl. 6); this specimen shows the presence of an M2, which despite the buccal wall being broken, is large and shows a subquadrated occlusal shape, in which the major axis is buccolingual, as is characteristic of Y. americana . Crassidia intermedia appears in our phylogenetic analysis ( Text-fig. 3 View Text-fig ) as the sister group of the rest of the Thaumastocyoninae , close to Y. gerandiana . Peignecyon felinoides differs from these two genera in a suite of advanced features, such as the loss of the m1 metaconid and the strong reduction of the M1 lingual area, which it shares with the most specialized Thaumastocyoninae genera, Tomocyon and Thaumastocyon .
Tomocyon grivensis from the late middle Miocene of La Grive ( France) differs from P. felinoides in the following characters: a p4 exhibiting a higher protoconid and a much stronger distal cingulum; m1 long, with a sharp mesial paraconidcristid, and buccolingually compressed, like the protoconid and talonid. The distolingual cristid of the protoconid is very prominent and has no metaconid, while the hypoconid has a strong basal cingulum extending distally beyond the distal edge of the tooth; in the m2 the metaconid is located distally to the very large and high protoconid, so that the trigonid valley is reduced and mesio-lingually located, the talonid is narrow and similar to what it is observed in the m1, the base of the hypoconid is distally expanded; the P4 is longer, with a strong parastyle and a more distal protocone; the M1 exhibits a wider lingual area; consequently, its occlusal shape is subquadrangular, the paracone is much higher and more developed than the metacone, the metaconule is poorly differentiated and the lingual cingulum is stronger, especially in the distolingual corner; in the m2 the protocone connects both the metaconule and the paraconule, forming a continuous semicircular crista, which joins the buccal cingulum. Many of these differences indicate that T. grivensis possesses a better developed sectorial dentition compared to that of P. felinoides . However, in the M1, it presents a larger lingual area in contrast to the reduction observed in the Thaumastocyon species ; this is also observed in the m2, which retains a relatively large size.
The other genus is Thaumastocyon , with two species, Th. bourgeoisi (type species) from the middle Miocene (MN 5) and Th. dirus from the late Miocene (MN 9) of Spain ( Ginsburg et al. 1981). As we have already pointed out, Thaumastocyon is characterized by extreme reduction of the M1 lingual cingulum feature, which is absent in all the other members of this subfamily. The holotype of Th. bourgeoisi STHELIN et HELBING, 1925 is a mandible (Pont- Levoy 69) preserving the canine and the m1–m2. The m1 has more compressed buccolingual cuspids than those of P. felinoides ; moreover, the reduction of the metaconid is complete, although a tiny notch in the distolingual cristid marks its ancestral position; the distobuccal base of the hypoconid is more swollen and projected distally. The m2 is morphologically similar to the morphotype, of Tuchořice, which presents a reduced talonid. Furthermore, Thaumastocyon dirus is somewhat larger, and differs from the type species of the genus in the almost total loss of the M1 lingual cingulum.
Ysengrinia valentiana from the locality of Buñol ( Spain), early Miocene (MN 4) ( Belinchón and Morales 1989) is quite interesting, although the species is represented by only two teeth: a left M1 (smaller in size that the M1 of P. felinoides ), which displays a reduced lingual area, with a weak lingual cingulum distally placed. The protocone, although still in a central position, presents a mesial crista that forms part of the mesial wall, similar to that of Y. americana . Additionally, the reduction of the labial cingulum, with the consequent buccolingual shortening of the molar, clearly differentiates Y. valentiana from the American species. The second tooth of Y. valentiana , left m2, is morphologically close to that of P. felinoides (mandible TU 73917; Pl. 2, Fig. 1); it presents a strong wear facet in the mesiobuccal corner, which perfectly fits with a wear facet on the lingual wall of the M1 metacone. In Y. valentiana , the molar morphology is close to Thaumastocyon bourgeoisi , but the Spanish species still retains a larger M1 lingual cingulum. At least regarding the M1, Y. valentiana certainly appears to occupy an intermediate position between P. felinoides and Th. bourgeoisi , although we have very little material to ascertain its systematic position.
| TU |
Tulane University, Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Peignecyon felinoides
| Morales, Jorge, Fejfar, Oldřich, Heizmann, Elmar, Wagner, Jan, Valenciano, Alberto & Abella, Juan 2019 |
Peignecyon felinoides
| Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella 2019 |
P. felinoides
| Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella 2019 |
P. felinoides
| Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella 2019 |
P. felinoides
| Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella 2019 |
P. felinoides
| Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella 2019 |
P. felinoides
| Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella 2019 |
P. felinoides
| Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella 2019 |
Ysengrinia
| GINSBURG 1966 |
Ysengrinia
| GINSBURG 1966 |
Amphicyon
| Lartet 1836 |