
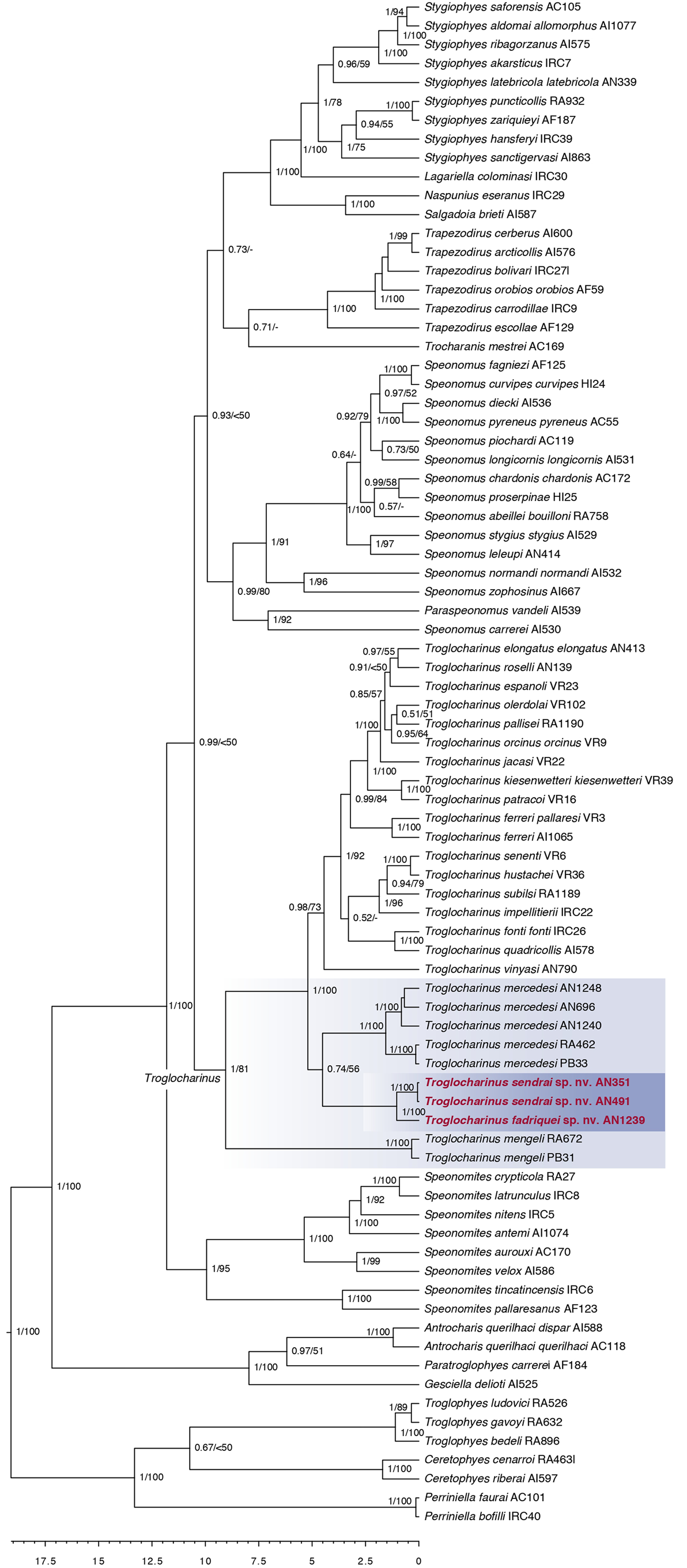
Troglocharinus Reitter, 1908
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4926.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:740D55C2-35EA-4D83-98D0-AC924C38CDD5 |
|
DOI |
https://doi.org/10.5281/zenodo.4534246 |
|
persistent identifier |
https://treatment.plazi.org/id/03E587FA-FFAB-FFAE-FF2A-5B052E7F7733 |
|
treatment provided by |
Plazi |
|
scientific name |
Troglocharinus Reitter, 1908 |
| status |
|
Troglocharinus Reitter, 1908 View in CoL
Perrinia Reitter, 1885: 16 ( Perrinia Adams, 1853 , Mollusca, Trochidae View in CoL )
Troglophyes (Troglocharinus) Reitter, 1908: 116
Troglocharinus Reitter View in CoL : Jeannel, 1910: 34
Speophilus Jeannel, 1911: 93 View in CoL
Antrocharidius Jeannel, 1910: 283 View in CoL
Type species: Troglophyes (Troglocharinus) ferreri Reitter, 1908
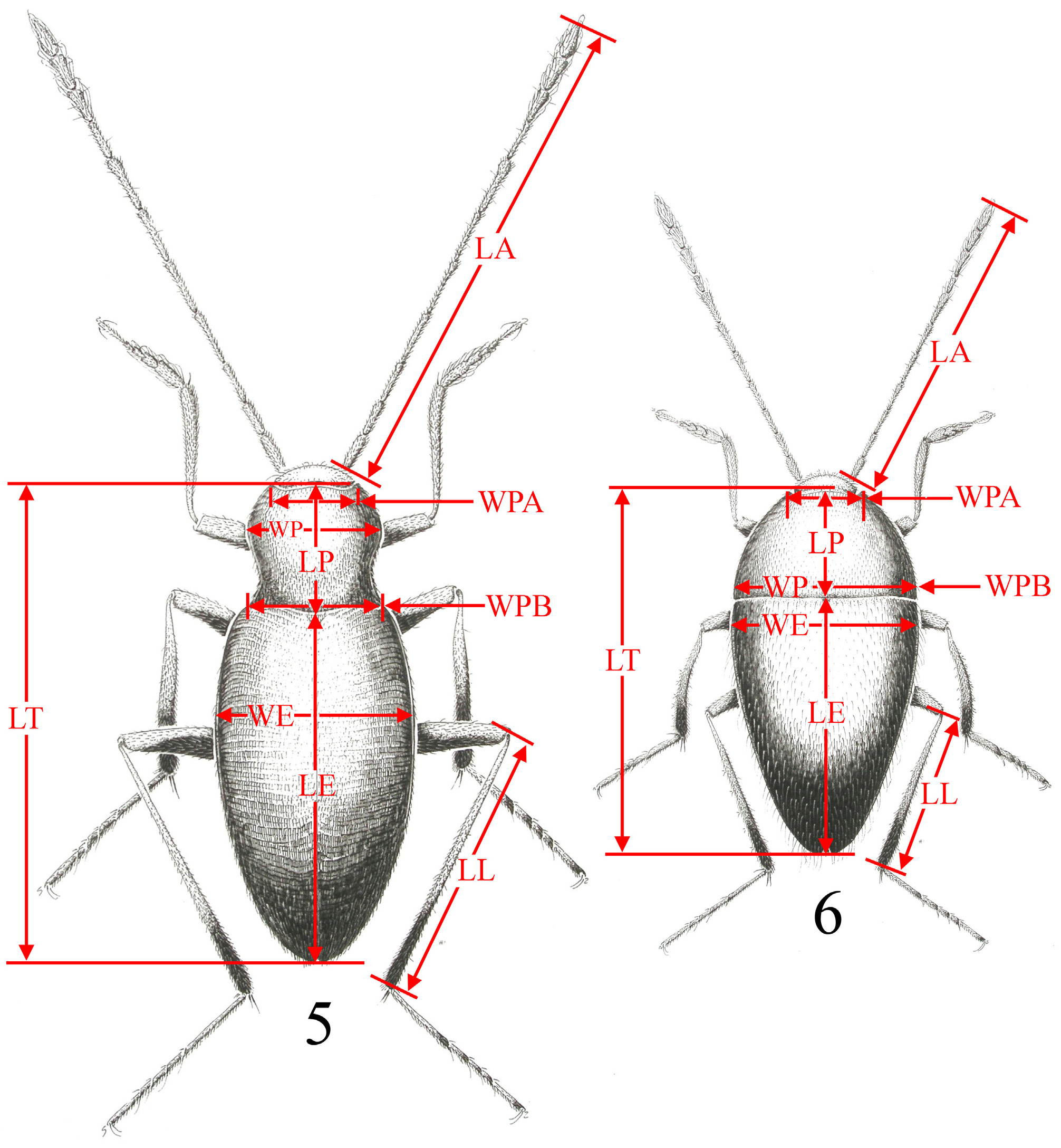
Species of Troglocharinus are hypogean, anophthalmous, apterous and depigmented, with a pholeuonoid (i.e. pronotum bell-shaped) or bathyscioid (i.e. pronotum trapezoidal) habitus ( Figs 3–5 View FIGURES 3–4 View FIGURES 5–6 ). Size ranging between 2.5–4.5 mm, with a yellowish-reddish general colouration; pubescence yellowish, fine, dense and recumbent, although in some cases with longer erect setae, mostly in the apical area of the elytra (as e.g. in T. patracoi (Zariquiey, 1922)) ; with irregular punctation on the pronotum and arranged transversally in the elytra, forming transverse striolas.
Antennae longer than the body, particularly in the species with more developed pholeuonoid habitus. Antennomeres i–vi slender, vii with engrossed apex, viii slender and ix-xi enlarged ( Table3 View TABLE 3 ).
Pronotum narrowing posteriad in some species ( T. ferreri , T. hustachei Jeannel, 1911 , T. impellitierii Español, 1955 and T. senenti Escolà, 1967 ), with sinusoid lateral margins ( T. elongatus Zariquiey, 1950 , T. espanoli (Jeannel, 1930) , T. fonti ( Jeannel, 1910) , T. jacasi (Lagar, 1966) , T. kiesenwetteri (Dieck, 1869) , T. ludovici Bellés & Déliot, 1983 , T. olerdolai Lagar, 1952 , T. orcinus ( Jeannel, 1910) , T. pallisei Rizzo & Comas, 2015 , T. patracoi (Zariquiey, 1922) , T. quadricollis ( Jeannel, 1911) , T. schibii (Español, 1972) and T. subilsi ( Español, 1965)) or regularly arched ( T. vinyasi ( Escolà, 1971)) .
Elytra long and narrow in pholeuonoid species, shorter and wider in bathyscioid species.
Mesoventral keel well developed ( Fig. 12 View FIGURES 12–14 ; T. fonti , T. kiesenwetteri , T. ludovici , T. quadricollis , T. senenti , T. subilsi and T. vinyasi ) or reduced ( Figs 13–14 View FIGURES 12–14 ; T. elongatus , T. espanoli , T. ferreri , T. hustachei , T. impellitierii , T. jacasi , T. kiesenwetteri andresi (Escolà 1966) , T. olerdolai , T. orcinus , T. pallisei , T. patracoi and T. schibii ).
Legs exceptionally long, in particular the metafemur of the more pholeuonoid species. Males with dilated first three protarsomeres; females with non dilated protarsomeres.
Female urite viii with short spiculum, with apex narrowly rounded ( Figs 58, 61, 62 View FIGURES 58–63 ) or, more frequently, bifurcated ( Figs 59–60 View FIGURES 58–63 ). The related genera from the Catalonian Pyrenees Pallaresiella and Speonomites have similar bifurcated urite viii ( Fig. 63 View FIGURES 58–63 ).
Metatergal apparatus very reduced ( Figs 68, 69 View FIGURES 64–69 ), with narrow and straight metascutum (SC3) (sensu Friedrich & Beutel 2006), without sutures; alacrista ( ALC) without apical node and very short basal apophisis (A).
Metendosternite with two long symmetric lateral arms, very narrow and diverging; stalk also narrow and as long as the arms ( Figs 64–66 View FIGURES 64–69 ). In Speonomites ( Fig. 67 View FIGURES 64–69 ) the stalk is ca. half the length of the arms. In the species transferred to Troglocharinus and the newly described ones the stalk is longer than in Speonomites , but still clearly shorter than the arms.
Median lobe of the aedeagus in lateral view strongly flexed in an obtuse or right angle between the middle part and the apical third ( Figs 15–19 View FIGURES 15–19 ); narrowing progressively from base to apex, which is pointed with perhaps a slight preapical dilatation; the dorsal part of the aedeagus may have a depression in the apical third with a very narrow extended apex; the sternal part of the apical third may be somewhat sinuate.
Lateral stylus of the tegmen (i.e. parameres) shorter than the median lobe ( Figs 15–24 View FIGURES 15–19 View FIGURES 20–24 ); with two setae inserted apically and one pre-apically, the first two long, robust and either straight, arched, slightly sinuous or abruptly curved in different sections; the third finer and as long as the others or much shorter and slightly sinuous or almost straight. With a well developed membranous lamina ( Figs 35–43 View FIGURES 35–47 ) or short and trapezoidal (e.g. in T. orcinus and subspecies and in T. pallisei Rizzo & Comas 2015 ; Fig. 44 View FIGURES 35–47 ), a derived state according to our phylogenetic results ( Fig. 1 View FIGURE 1 ); with a tuf of long hairs (“penicillum”) in the surface of the lamina, especially along the margins, absent only in T. orcinus and subspecies and in T. pallisei ( Rizzo & Comas 2015; Fig. 44 View FIGURES 35–47 ). The length of the lamina and the tuff of setae seem to be related, as both characters always occur together. With some sensilla placoidea close to the apex.
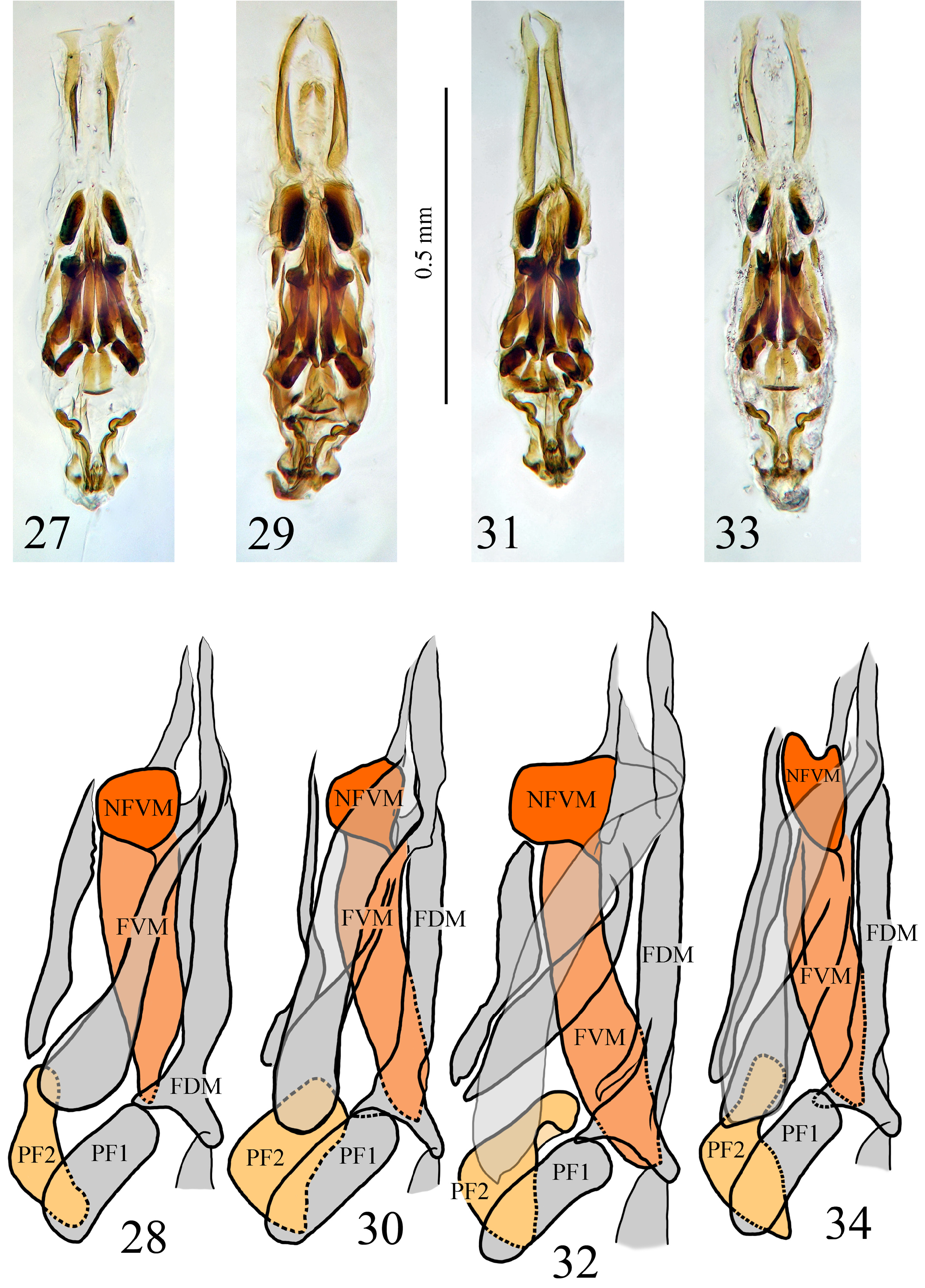
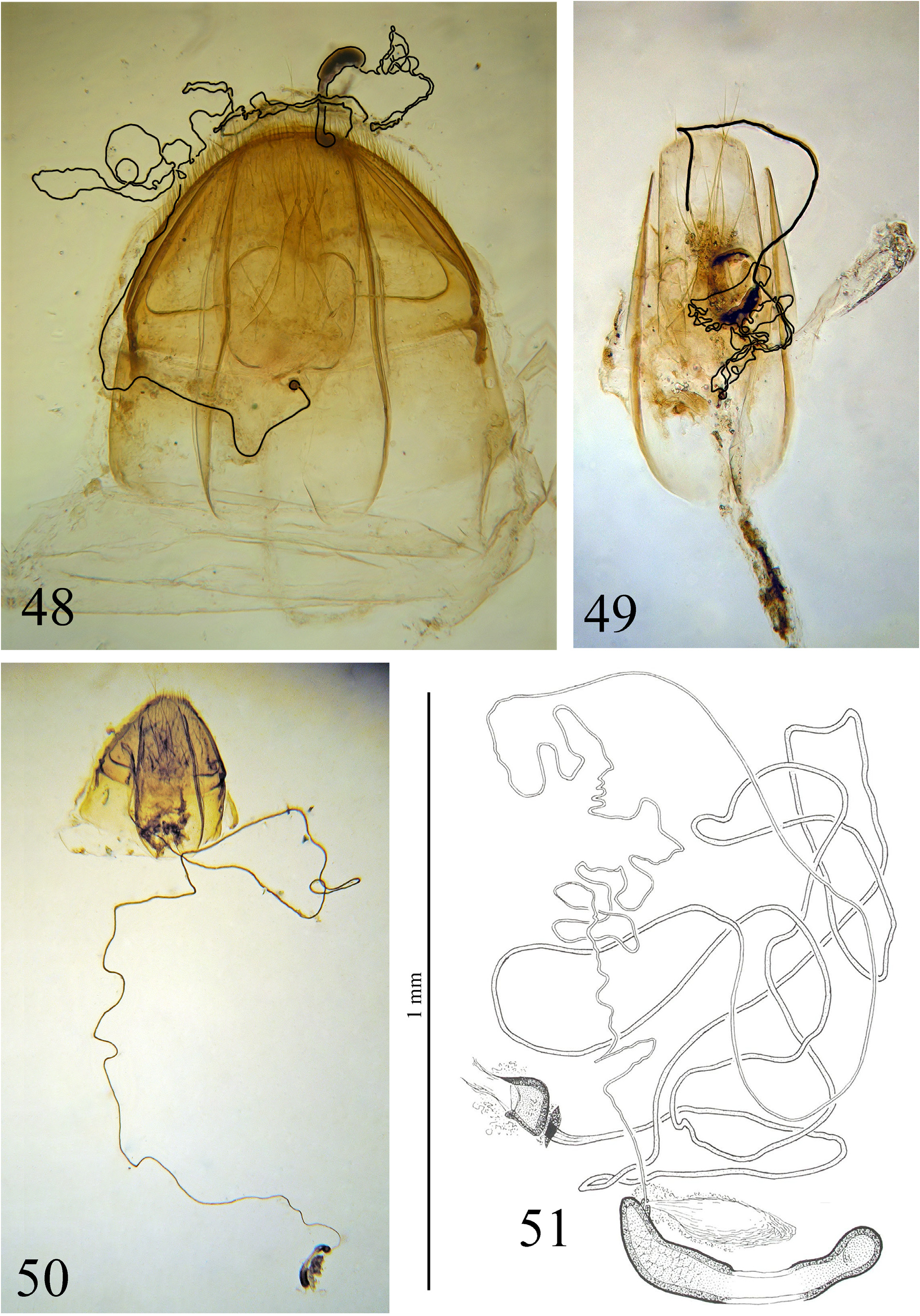
Endophallus with three sections as defined by Jeannel (1924) ( Figs 27–34 View FIGURES 27–34 ). Basal region (RB) with an “Ypiece” (PY) with divergent and sinuous arms, articulated with the apical membranous extension through a nodule; these structures form a funnel with the floating pieces and the appendices of the dorsal feather-like structures of the medial region ( FDM) in the upper part. Medial region with two enlarged feather-like structures, bifurcated at the base; one of the bifurcations extented until the funnel-like structure of the basal region; with a floating piece in between the two arms of the bifurcated feather-like structures also in contact with the funnel-like structure. With two ventral feather like structures ( FVM) enlarged and divergent at their base, with a nodule of variable shape in the apex; with two upper dorsal plates, each of them with a long and thin external, and another more robust internal, feather-like structures, the later with the apex bifurcated and the enlarged area folded in a different orientation, very sclerotised. With two large bags at both sides of the axial piece, with very sclerotised internal nodules ( NC) and a long appendix at the base. Apical region with reinforcement bands ( BRA) with a bag in between them, with longitudinal stripes and the margins engrossed.
Female genitalia ( Figs 52–57 View FIGURES 52–57 ) with an elongated spermatheca, with two well sclerotised lobes and a hyaline medial region, narrower than the terminal lobes; apical lobe hemispherical. Spermaduct and spermatic gland inserted in the basal region; spermatic duct between 40–70 times the length of the spermatheca, with two sections of different diameter and a basal process in the bursa copulatrix ( Fig. 51 View FIGURES 48–51 ).
The presence of a large membranous lamina (in comparison to other genera with a smaller lamina, as e.g. Speonomites , Figs 45–47 View FIGURES 35–47 ) with a tuff of setae in the apex of the lateral stylus of the median lobe (the “penicillum”), and the very long spermatic duct with two sections with a different diameter and a basal process in the bursa copulatrix are two potential synapomorphies of the genus Troglocharinus . The first, presence of a large lamina with “penicillum” in the apex of the parameres, is secondarily lost in the species T. orcinus and its subspecies ( Fig.44 View FIGURES 35–47 ), although some populations may present intermediate degrees of development of this structure (unpublished observations). There are two further apomorphies of the genus shared by related genera: antennae longer than the body, present also in Speocharidius Jeannel, 1919 from the Vasc Country and other Leptodirini of different faunas, and very long legs, specially the metafemora, also present in Speocharidius and other Leptodirini.
Both synapomorphies are present in the species formerly in Speonomites transferred here to Troglocharinus ( T. mengeli and T. mercedesi ) and the two newly described species (see below). These four species have a bathyscioid habitus, characterised by a rather rounded body form and shorter appendages and antennae ( Figs 3–4 View FIGURES 3–4 ), compared with the pholeuonid habitus of other species, and a well developed mesoventral keel.
The phylogenetic reconstruction ( Fig 1 View FIGURE 1 ) shows that Pallaresiella pallaresana ( Jeannel, 1911) —code voucher AF123—is the sister species of Speonomites tincatincensis (Escolà, Bellés & Comas, 1985) —code voucher IRC6— with a support value ( ML) of 100, and both are sister of the genus Speonomites Jeannel, 1910 . Since the structure of the endophallus is similar and the two genera present the same distribution area, we establish the synonymy between the genus Speonomites and Pallaresiella Fresneda, 1998 syn. nv.







| BRA |
Slovak National Museum |
| ML |
Musee de Lectoure |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Troglocharinus Reitter, 1908
| Fresneda, Javier, Rizzo, Valeria, Comas, Jordi & Ribera, Ignacio 2021 |
Speophilus
| Jeannel, R. 1911: 93 |
Troglocharinus
| Jeannel, R. 1910: 34 |
Antrocharidius
| Jeannel, R. 1910: 283 |
Troglophyes (Troglocharinus)
| Reitter, E. 1908: 116 |
Perrinia
| Reitter, E. 1885: 16 |