
Nemertopsis quadripunctata ( Quoy & Gaimard, 1833 )
|
publication ID |
https://doi.org/10.5281/zenodo.176154 |
|
DOI |
https://doi.org/10.5281/zenodo.6246623 |
|
persistent identifier |
https://treatment.plazi.org/id/03E32426-FFD4-8E22-FF45-52E2ADB6E4CD |
|
treatment provided by |
Plazi |
|
scientific name |
Nemertopsis quadripunctata ( Quoy & Gaimard, 1833 ) |
| status |
|
Nemertopsis quadripunctata ( Quoy & Gaimard, 1833) ( Figs 2–6 View FIGURE 2 View FIGURE 3 )
Borlasia quadripunctata Quoy & Gaimard, 1833 View in CoL Colpocephalus quadripunctatus: Diesing, 1850 Nemertopsis gracilis: Iwata, 1954
Nemertopsis quadripunctatus [sic]: Gibson, 1990
Material examined: Twenty live individuals, intertidal, among the gooseneck barnacle, Capitulum mitella (Cirripedia: Lepadomorpha), Shirahama, Wakayama Prefecture, Japan, 8 July 1999; ZIHU-3207, serial transverse sections of the anterior tip of the head (about 1 cm long), 6 µm, 15 slides.
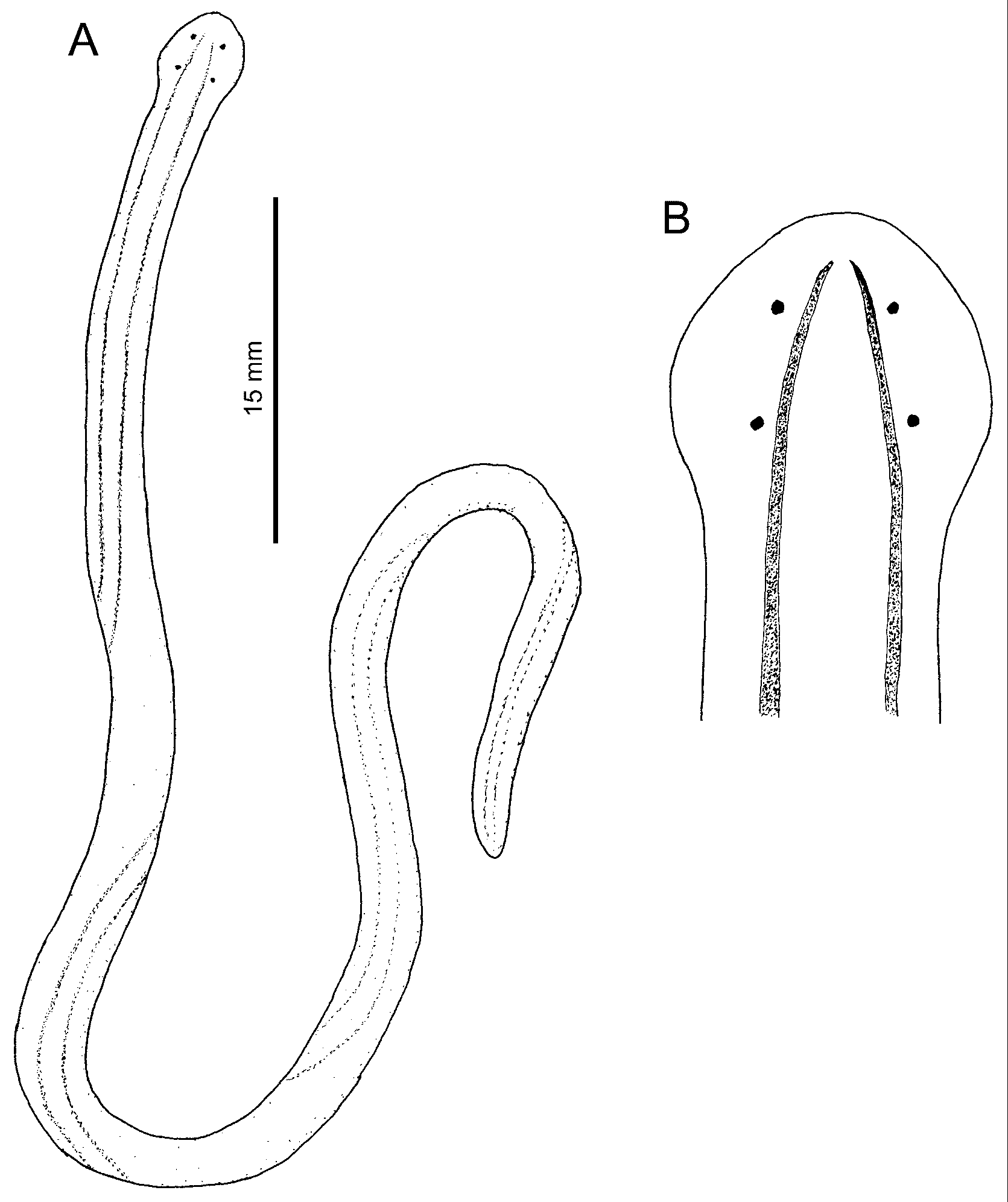
External features: Specimens examined alive ranged in length from 5–8 cm, in width from 0.6–1.0 mm in normal creeping state before anaesthetization. Body slender, rather voluminous, gradually narrowing near posterior tip of body, ending in bluntly pointed tail ( Fig. 1A View FIGURE 1. A – D ). Head rounded, wider than succeeding body, possessing four eyes arranged in corners of trapezium ( Fig. 1B View FIGURE 1. A – D ). No evidence of cephalic furrows on dorsal surface. Body colour milky white, dorsally marked by two dark brown stripes not meeting anteriorly ( Fig. 1A, B View FIGURE 1. A – D ).
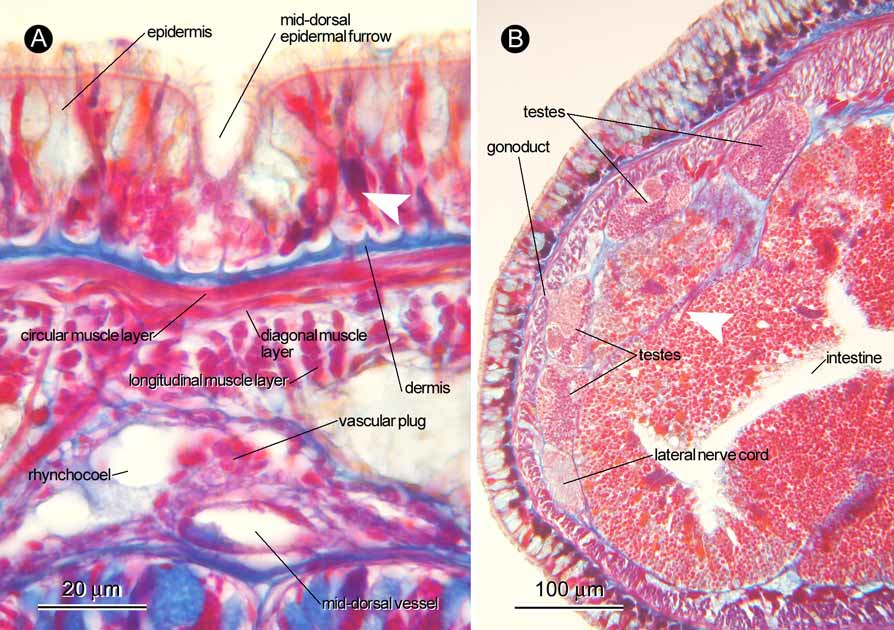
Body wall, musculature, and parenchyma: Ciliated epidermis ( Fig. 3 View FIGURE 3 A), mostly 30–40 µm thick in brain region, possessing strongly acidophilic glandular cells arranged proximally in post-cerebral region of body. Mid-dorsal epidermal furrow present ( Fig. 3 View FIGURE 3 A), extending from tip of head above apical organ, pre-cerebrally interrupted three times, post-cerebrally continuous at least to intestinal region; strongly acidophilic glandular cells absent in proximal portion of furrow ( Fig. 3 View FIGURE 3 A). Dermis, mostly 2–3 µm thick, its distal surface forming cup-like structure ( Fig. 3 View FIGURE 3 A). Body wall musculature composed of outer circular and inner longitudinal muscle layers; diagonal muscle layer of lattice type present between outer circular and inner longitudinal muscle layers ( Fig. 3 View FIGURE 3 A). In foregut region, outer circular layer up to 2–3 µm thick, inner longitudinal layer 8–30 µm thick. Dorsoventral muscle bundles found in foregut and intestinal regions ( Figs 3 View FIGURE 3 B, 4D). In foregut region, ventro-lateral portions of body-wall inner longitudinal muscle layer sending fibres medio-anteriorly traversing through basophilic sub-muscular glands (= posterior continuation of cephalic glands) ( Fig. 4 A, B); these fibres running forward immediately adjacent to ventro-lateral side of lateral nerve to form muscular plate ( Fig. 4 C, D); in brain region, muscular plate on each side of body uniting between ventral cerebral commissure and oesophagus ( Fig. 4 E, F); anterior to brain ring, muscle plate latero-dorsally extending upward along edge of brain to loosely surround anterior surface of brain ( Fig. 4 F, G), fibres from muscular plate eventually contributing to proboscis insertion ( Fig. 4 G, H). Additional fibres from body-wall longitudinal muscle layer in brain region also appears to contribute to proboscis insertion ( Fig. 4 G). Most fibres from muscle plate terminating at proboscis insertion, but small amount of fibres from muscle plate leading farther anteriorly beyond proboscis insertion, running around rhynchodaeum and oesophagus ( Fig. 4 H). Pre-cerebrally, muscle fibres from body-wall inner longitudinal muscle layer running criss-cross. Parenchymatous connective tissues not well developed.
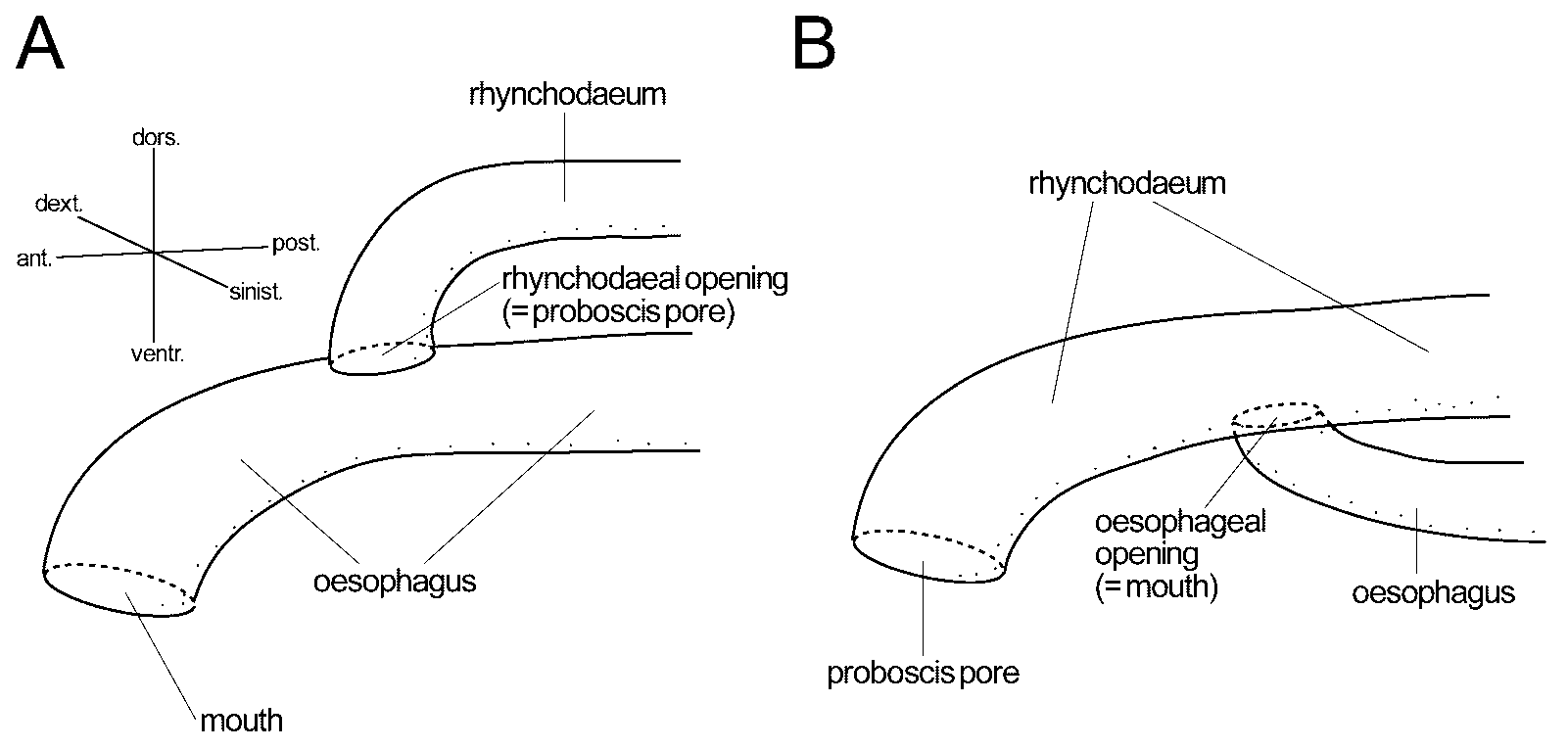
Proboscis apparatus: Rhynchodaeum opening into dorsal wall of oesophagus ( Figs 5 D, 6A), with epithelium mostly about 5 µm thick, possessing neither cilia nor glandular cells ( Fig. 5 E, F). Rhynchodaeal sphincter not found. Rhynchocoel narrow and shorter than body length, with wall composed of thin muscle layer ( Figs 3 View FIGURE 3 A, 4E, 5G). Proboscis missing in sectioned material.
Alimentary system: Oesophagus opening subterminally ( Figs 5 B, 6A), basally invested with longitudinal muscle fibres, with epithelium lacking cilia and gland cells, up to 8 µm thick ( Fig. 5 C–F). Stomach histologically divisible into three regions: anterior and posterior portions possessing epithelium dominated by acidophilic glandular cells ( Fig. 5 G, I); middle portion mainly containing basophilic glandular cells ( Fig. 5 H); stomach epithelium mostly 20–50 µm thick. Pylorus less glandular, opening to dorsal wall of intestine ( Fig. 5 J). Intestinal caecum with neither anterior nor lateral diverticula, extending below stomach to reach middle portion of stomach ( Fig. 5 H, I). Main intestinal canal with lateral diverticula.
Blood vascular system: Pair of cephalic vessels meeting anteriorly above rhynchodaeal opening ( Fig. 5 E), posteriorly passing through cerebral ring ( Fig. 4 E, F), leading to lateral vessels ( Fig. 4 D). Mid-dorsal vessel branching off from right lateral vessel, entering rhynchocoel just posterior to dorsal ganglia to form vascular plug, 20 µm in diameter ( Fig. 3 View FIGURE 3 A); mid-dorsal vessel soon coming down to run between rhynchocoel and alimentary canal. No transverse metameric connectives between mid-dorsal and lateral vessels.
Nervous system: Brain with outer neurilemma, but with neither inner neurilemma nor neurochord cells. Dorsal and ventral commissures about 20 µm and 45 µm thick respectively ( Fig. 4 F). Lateral nerve cords with neither accessory lateral nerve nor neurochord, containing single myofibril running in upper part of neuropile ( Fig. 4 D).
Apical organ and cephalic glands: Single apical organ present ( Fig. 5 A), about 20 µm wide, opening at tip of head just anterior to oesophagus opening. Basophilic cephalic glandular lobes ( Fig. 5 B, C) extending post-cerebrally as sub-muscular glands beneath body-wall longitudinal muscle layer ( Fig. 4 A), enormously developed in foregut region filling space between body-wall longitudinal muscle layer and various internal organs, gradually reducing posteriorly to be sporadically situated above and below lateral nerve cords. Acidophilic sub-muscular glandular cells scattered throughout body among basophilic glands ( Fig. 4 C), especially dense in dorsolateral portion of foregut and ventral side of intestine. Numerous ducts leading ventrally from acidophilic glandular cells, passing through body wall to open in epidermis.
Sense organs: Pigment cup ocelli about 60–70 µm in diameter ( Fig. 5 C, F); anterior pair situated near tip of head, posterior pair just in front of proboscis insertion. Cerebral organs simple, about 50 µm in diameter, situated in front of brain ( Fig. 5 F); cerebral organ canal opening ventrally near anterior eyes ( Fig. 5 C).
Excretory system: Excretory system extending from brain region to anterior intestinal region, consisting of thin-walled collecting tubules, up to 20 µm in diameter, running along lateral nerve cords in foregut region ( Fig. 4 B); efferent canals opening laterally or ventrally to exterior ( Fig. 4 A, B); two nephridiopores on left side, one on right.
Reproductive system: The sectioned material was a mature male. Testes distributed around lateral margins of body above lateral nerve cords between intestine and body-wall longitudinal muscle layer ( Fig. 3 View FIGURE 3 B).
Remarks: As to the external features, specimens from Shirahama differ from those of Ambon and Hong Kong in that their dorsal body coloration between the two longitudinal stripes is the same as that outside the stripes ( Iwata 1954; present study) rather than possessing a light yellowish tinge as reported by Quoy and Gaimard (1833) and Gibson (1990). In addition, Gibson (1990) noted that the dorsal stripes usually met at the tip of the head in Hong Kong material, whereas in the present material, as well as in the previous descriptions by Quoy and Gaimard (1833) and Iwata (1954), these did not unite anteriorly. The body length observed in the present study measured 5–8 cm, while Iwata (1954) recorded it varied from 10–12 cm in specimens collected in Shirahama, the same locality as the present study. Whether this difference in body size is due to seasonal variety or age remains uncertain, since Iwata (1954) did not give the date of collection of his samples. By comparison, Gibson’s (1990) material from Hong Kong ranged from 1.5–61 mm, though he quoted student reports giving a maximum length of 80 mm. Quoy and Gaimard (1833) noted the worm was “a tout au plus deux pouces de longeur” (= at most two inches in length = about 5 cm).
The mid-dorsal epidermal furrow, an unusual feature among whole phylum and first described by Gibson (1990) [‘dorsal epidermal pit’ in his terminology], was also confirmed in the present material. In Hong Kong specimens, the pits are reported to be arranged in a row ( Gibson 1990). In the present specimen from Shirahama, the ‘pits’ are antero-posteriorly continuous to a large degree, forming a sagittal furrow along the middorsal portion of the epidermis. The furrow is discontinuous by three short shallow portions in the pre-cerebral region, and completely contiguous post-cerebrally, at least in the sectioned part of the body (anterior tip, 1 cm long).
Gibson (1990) noted that a mid-dorsal vascular plug was not found in Hong Kong specimens, probably due to the small size of the body; the vascular plug may difficult to observe when it is pressed by the proboscis against the ventral surface of the rhynchocoel. The sectioned specimen from Shirahama had lost its proboscis, which enabled confirmation of the presence of the vascular plug, in spite of the narrowness of the rhynchocoel in the brain region ( Fig. 3 View FIGURE 3 A).
Unlike in most monostiliferous hoplonemerteans, the rhynchodaeum in the present material appeared to open to the dorsal portion of the oesophagus ( Fig. 6 View FIGURE 6 A); the canal anterior to the oesophagus-rhynchodaeum junction appeared posteriorly continuous to the oesophagus, instead of the rhynchodaeum, both as to the width of the canal and the nature of its wall, including the thickness and histological appearance ( Fig. 5 C–E). A similar oesophageal-rhynchodaeal relationship has been recorded in five species of monostiliferous hoplonemerteans: Africanemertes ruetzleri Kirsteuer, 1965 ; Africanemertes swakopmundi Stiasny-Wijnhoff, 1942 ; Divanella evelinae Gibson, 1973 ; Itanemertes nonatoi Corrêa, 1957 ; and Nemertopsis bivittata ( Delle Chiaje, 1841) sensu Corrêa (1955) . Meanwhile, specimens of N. quadripunctata from Hong Kong are reported to possess a normal oesophagus-rhynchodaeum configuration ( Fig. 6 View FIGURE 6 B), i.e., oesophagus opening to ventral portion of rhynchodaeum ( Gibson 1990). Future studies will elucidate the intra- and inter-populational variation of this character.
In life, the worms secreted viscous mucus from their ventral surface; the viscosity of the mucus was so strong that when an attempt was made to peel a worm away from the surface of a petri dish by sucking up a free end with a pipette, the animal was torn in two.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Nemertopsis quadripunctata ( Quoy & Gaimard, 1833 )
| Kajihara, Hiroshi 2007 |
Nemertopsis gracilis :
| Iwata 1954 |
Colpocephalus quadripunctatus :
| Diesing 1850 |
Borlasia quadripunctata
| Quoy & Gaimard 1833 |