
Adenomera phonotriccus, De Carvalho & Giaretta & Angulo & Haddad & Peloso, 2019
|
publication ID |
https://doi.org/10.1206/3919.1 |
|
DOI |
https://doi.org/10.5281/zenodo.4585379 |
|
persistent identifier |
https://treatment.plazi.org/id/03E287D8-FFDF-0250-49C6-DA49FC57C3C7 |
|
treatment provided by |
Felipe |
|
scientific name |
Adenomera phonotriccus |
| status |
sp. nov. |
Adenomera phonotriccus View in CoL , n. sp.
Figures 1‒3 View FIG View FIG View FIG
REFERRED MATERIAL: Voucher specimens with DNA sequenced by Fouquet et al. (2014) and assigned to Adenomera sp. F (in part).
SUGGESTED VERNACULAR NAME: Tody-tyrant-voiced nest-building frog.
HOLOTYPE: MPEG 41155 View Materials (field number TRC 135 ), adult male, from the municipality of Palestina do Pará ( 5.70228° S, 48.24949° W; 166 m), on the western margin of the lower Araguaia River , state of Pará, northern Brazil, collected by P.L. V. Peloso, M.J. Sturaro, P. V. Cerqueira, G. Gonçalves, A.A. Giaretta, P. Marinho, and T. R. de Carvalho on January 9, 2018 (Zoobank LSID registration: urn:lsid:zoobank.org:act:). GoogleMaps
SOUND RECORDING: Adenomera _phonotriccusPalestinaPA2bAAGm661 MK 2.
PARATOPOTYPES: MPEG 41156 View Materials and CFBH 43130‒43131 View Materials , all adult males, collected ( 5.70300° S, 48.24833° W; 176 m) on January 9‒10, 2018, by the same collectors GoogleMaps .
ETYMOLOGY: The epithet phonotriccus is the combination of Greek phono - (from phoné, “sound, voice”) and triccus (a small bird whose species is not identifiable, though in modern times the name is applied to tyrant flycatchers; Jobling, 2010). This name is to be treated as a noun in apposition and is an allusion to the similarity between the vocalization of the new species and those of tody-tyrants. Tody-tyrants of the Neotropical genus Hemitriccus have peculiar vocalizations, which are reminiscent of the vocalization of Adenomera phonotriccus , especially the trilled song of H. cohnhafli (see Zimmer et al., 2013). Interestingly, several Hemitriccus species are morphologically very similar, and consequently best differentiated by their vocalizations, a pattern also observed in Adenomera .
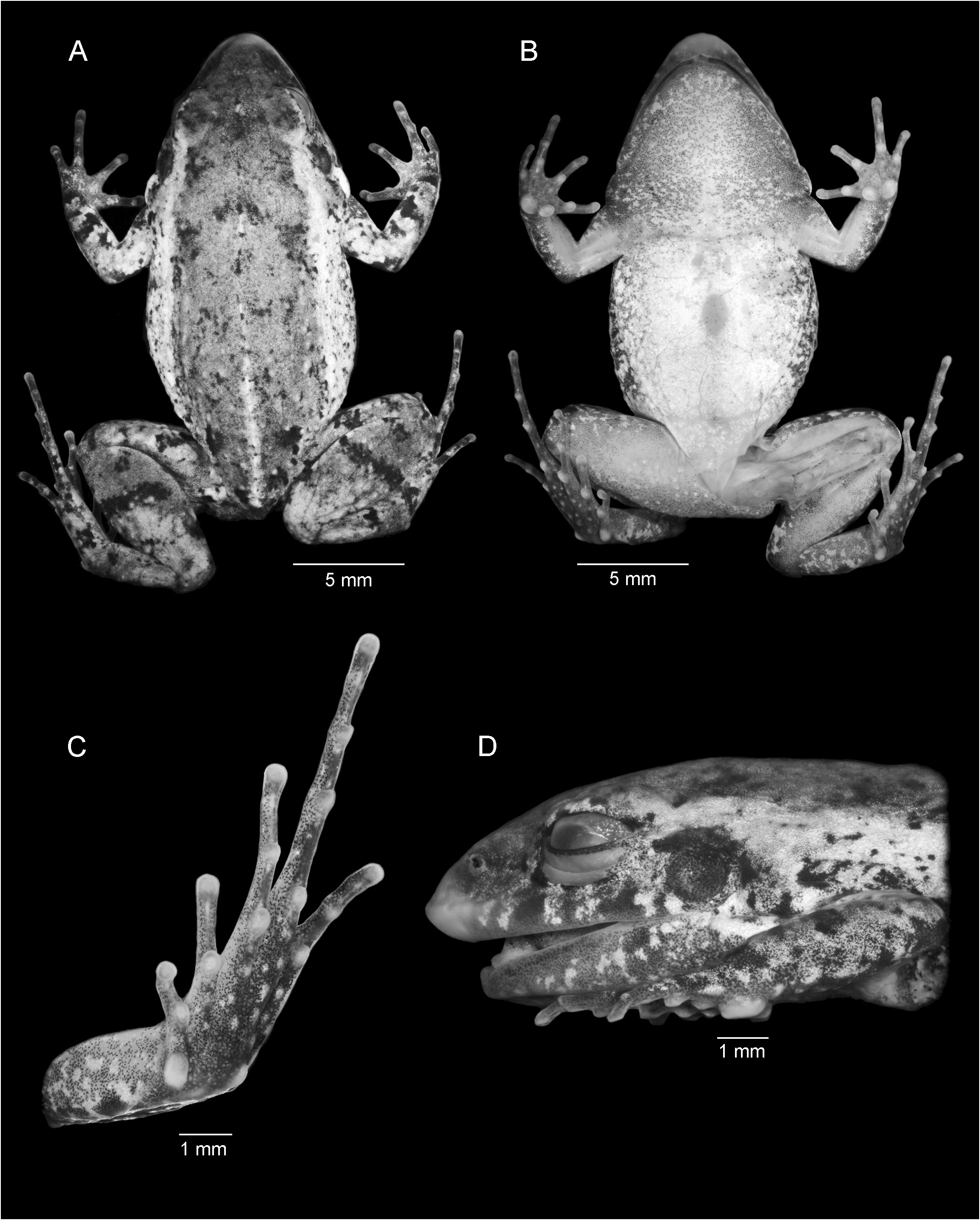
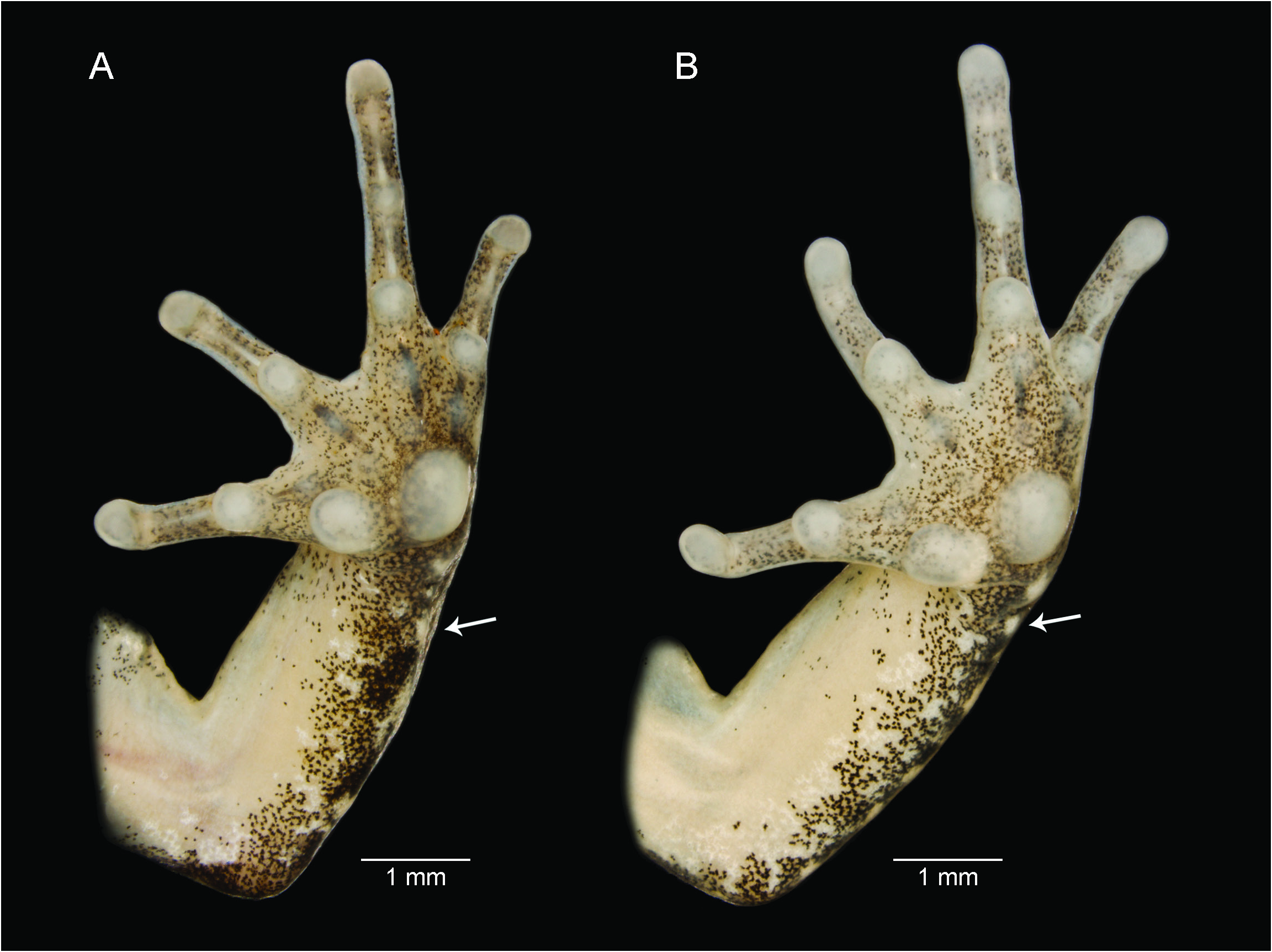
DIAGNOSIS: Adenomera phonotriccus is differentiated from its congeners by the following combination of characters: (1) small size (adult male SVL 19.8–21.6 mm; table 1); (2) robust body shape; (3) toe tips unexpanded to slightly swollen (character states B, C; Heyer, 1973); (4) distal antebrachial tubercle on underside of forearm; (5) throat and belly cream colored, mottled white and gray in some parts; (6) two color morphotypes (presence/absence of dorsolateral stripes); (7) advertisement call consisting of a single type of pulsed note, emitted regularly, not in calling bouts; (8) advertisement notes composed of complete pulses (pulses with periods of silence in between); (9) long-lasting call duration (213‒433 ms).
COMPARISONS: Adenomera phonotriccus has adult males (SVL 19.8–21.6 mm; table 1) smaller than those of A. coca (23.6–25.6 mm; Angulo and Reichle, 2008), A. lutzi (25.7–33.5 mm; Kok et al., 2007), and A. simonstuarti (SVL 25.9–26.2 mm; Angulo and Icochea, 2010). Adenomera phonotriccus has a robust body shape (figs. 1–2), whereas A. diptyx , A. martinezi , and A. saci have a slender body ( Carvalho and Giaretta, 2013a). Adenomera phonotriccus has toe tips unexpanded or slightly swollen (character states B, C), but not expanded into flattened discs (character state D) as in A. ajurauna , A. andreae , A. marmorata , A. lutzi , A. nana , and A. simonstuarti ( Angulo et al., 2003; Kok et al., 2007; Kwet, 2007; Berneck et al., 2008; Angulo and Icochea, 2010; appendix 1). Adenomera phonotriccus is distinguished from congeners (except A. cotuba and A. lutzi ; Kok et al., 2007; Carvalho and Giaretta, 2013b) by having antebrachial tubercles on underside of forearm (fig. 3), presumably absent in all remaining species. Adenomera phonotriccus differs from A. heyeri and A. lutzi by having ventral surfaces cream colored (mottled white and gray in some parts), yellow in the latter two species ( Boistel et al., 2006; Kok et al., 2007). From A. engelsi , which does not have distinctive dorsolateral stripes and pigmentation on throat ( Kwet et al., 2009), Adenomera phonotriccus differs by the occurrence of both color features (fig. 1). Adenomera cotuba does not have dorsolateral stripes ( Carvalho and Giaretta, 2013b), whereas A. phonotriccus has two color morphotypes (presence/ absence of dorsolateral stripes). These species are better distinguished by their distinct vocalizations (see below; fig. 4).
The advertisement call of Adenomera phonotriccu s (fig. 4; table 2) distinguishes it from all congeners by having pulsed notes composed of complete pulses (periods of silence in between), whereas the other species’ notes are formed by incomplete pulses, i.e., not separated one from the next by periods of silence (fig. 4). From the remaining species of Adenomera , the new spe- cies differs by having pulsed advertisement notes, whereas those species’ calls consist of nonpulsed notes (table 3). The advertisement call of A. phonotriccus is distinguished from that of A. cotuba by being composed of single notes emitted regularly (table 2), whereas A. cotuba’ s call is made up of many notes emitted as calling bouts, i.e., multinote advertisement call ( Carvalho and Giaretta, 2013b: fig. 4A). In addition, to date, Adenomera phonotriccus has the longest note duration in the genus, whose range does not overlap with note-duration values for any congeners (table 3).
DESCRIPTION OF HOLOTYPE: MPEG 41155 (figs. 1A, 2, 3A). Adult male. Body robust (figs. 1A, 2A). Snout subovoid in dorsal view (fig. 2A), acuminate in lateral view (fig. 2D). Nostrils closer to the snout tip than to the eyes; fleshy ridge on snout tip; canthus rostralis not marked; loreal region slightly concave; supratympanic fold from the posterior corner of the eye to the base of the arm; oval postcommissural gland; vocal sac subgular with a fold from jaw extending to forearm on each side, vocal slits present; vomerine teeth in two straight rows medial and posterior to choanae and oblique to sagittal plane. Tongue elongated, free behind. Relative finger lengths IV <I <II <III; fingers without ridges or fringes; finger tips rounded, slightly expanded, but not flattened; inner metacarpal tubercle nearly rounded; outer metacarpal tuber- cle rounded (fig. 3A). Subarticular tubercles rounded; supernumerary tubercles discrete, rounded. Antebrachial tubercle on distal end of forearm. Dorsum shagreened, warty on flanks and inguinal region. Throat and center of belly smooth (fig. 2B). Belly granular laterally and posteriorly. A pair of lumbar glands on sacral region. Ventral surface of thighs granular. Posterior surface of thighs with no distinctive pattern, possessing discrete and rounded paracloacal glands. Relative toe lengths I <II <V <III <IV; toes without ridges or fringes; toe tips rounded and slightly swollen in toes II–IV. Inner metatarsal tubercle oval, outer metatarsal tubercle rounded (fig. 3C). Tarsal fold from the inner metatarsal tubercle extending 1/2 length of tarsus. Subarticular tubercles rounded or subconical; supernumerary tubercles rounded, discrete. Measurements of the holotype are provided in table 1.
COLOR OF HOLOTYPE IN PRESERVATIVE (fig. 2): Snout tip with a faded white coloration (coincident with the fleshy ridge). Dorsum marble gray with a few darker specks and spots. Sacral stripe on the posterior third of body length, cream-colored. Black lumbar glands. Dorsal surface of limbs with dark brown stripes/blotches on a slightly lighter brown background. Dorsolateral, broad stripe from the posterior corner of the eye, passing over the flanks, predominantly cream-colored to a somewhat orange hue (faded coloration; orange in life). Upper and lower jaws covered with white-colored spots/blotches, whitish postcommissural gland, outlined at the bottom by a dark coloration. Tympanum reddish brown. Throat, belly, and ventral surface of limbs cream, with melanophores, throat with a black-dotted pattern, solid dark-colored laterally, coinciding with the expanded vocal sac. Posterior surface of thighs with a few dark brown spots on a light brown background, and paracloacal glands cream colored.
COLOR OF HOLOTYPE IN LIFE (fig. 1A): Dorsum covered with black specks and spots irregularly distributed on medium brown background. Arms and legs reddish brown. Sacral and dorsolateral stripe, and postcommissural gland orange. Iris copper. Tympanum brown. Ventral surfaces cream colored and with mottling on throat and belly (laterally). Groin yellow.
VARIATION IN THE TYPE SERIES: Despite minor variation in dorsal coloration and distribution of spots, specks, and warts on dorsal surfaces, characteristics of the three paratypes largely agree with the description of the holotype. Dorsolateral stripe is absent in two paratypes ( MPEG 41156 View Materials and CFBH 43130 View Materials ) . Shape and size of antebrachial tubercle varies among the type series; some of them are very discrete (fig. 2).
ADVERTISEMENT CALL: The description is based on recordings of three males (appendix 3). Sample sizes for each acoustic trait and descriptive statistics, i.e., means and standard deviations, are provided in table 2. The call (fig. 4) consists of a single type of pulsed note, which is emitted at a rate of 26–35 calls per minute. Note duration varies from 213–433 msec. Notes have a smooth attack and decay, reaching the amplitude peak at 43%–85% of their duration. Pulse number varies from 14–26; pulse duration from 4–26 msec, emitted at a rate of 51–67 pulses per second; silence between pulses varies from nearly zero to 13 msec. The fundamental frequency of the note occupies a narrow bandwidth harmonic, with a peak frequency of 1857– 1997 Hz; the dominant frequency varies from 3639–4113 Hz and is contained within the second harmonic. Notes usually have a slight positive frequency modulation along their duration, but a few notes do not have any modulation, and a single note has negative modulation. Linear frequency modulation varies from -43–861 Hz.
HABITAT AND NATURAL HISTORY: Adenomera phonotriccus is associated with forest habitats in Brazil’s eastern Amazonia in the state of Pará. The type locality is located on the west margin of the lower Araguaia River. This region is also occupied by savanna landscapes or ecotones between the Cerrado and Amazonian rainforest. The species was heard along an open area transect (by recent logging activity), especially at the forest edge, and inside the forest remnant. Males called while exposed or under leaf litter, and increased their calling activity during and shortly after rainfalls. The vocalization of Adenomera phonotriccus is very peculiar when heard in the field. Our first impression was that the calls might not be produced by a leptodactylid frog, or even an anuran species. Indeed, the vocalization is similar to that of some Neotropical suboscine birds (e.g., Rhynchocyclidae , Tyrannidae ). There were two partially syntopic species of Adenomera also in calling activity at the study site: Adenomera sp. ( A. heyeri clade) and A. aff. hylaedactyla . We heard the three species calling at the same period (late afternoon), even though A. phonotriccus and Adenomera sp. ceased most calling activity at dusk, whereas A. aff. hylaedactyla continued calling through the first hours of the night. The last species occupied open areas, e.g. pasture. In contrast, A. phonotriccus and Adenomera sp., are associated with forest habitats.
DISTRIBUTION (fig. 5): Adenomera phonotriccus is known with certainty only from the type locality and Marabá. However, specimens from other regions in the state of Pará were also assigned to this lineage, referred as Adenomera sp. F, by Fouquet et al. (2014). Given the high levels of cryptic species diversity in Adenomera and complex genetic structure within the lineages identified by Fouquet et al. (2014), we conservatively restrict the species’ distribution to the type locality region until additional data, especially vocalizations, are obtained for the other populations assigned to Adenomera sp. F, a lineage widely distributed in central-east Pará, northern Brazil.
CONSERVATION STATUS: Because the species is apparently restricted to forested habitats, or its immediate margins (forest edge), we infer that the current population trend for this species is “declining.” Major threats are deforestation (destruction of habitats) due to conversion of large areas where the species occur into pasture and agricultural land. The species appears to be locally abundant (although difficult to observe), which means that as long as its habitat is preserved the species should have its future secured. The known range of the species is hard to determine with current evidence. It may be much wider, if all the clade F specimens refer to A. phonotriccus , in which case the species would be found from the western margin of the Tocantins River to the western margins of the Tapajós River ( Fouquet et al., 2014: fig. 2). Alternatively, it could have a much narrower distribution (southern Pará) if only specimens in subclade F3 correspond to A. phonotriccus —in the latter case, the species would be probably restricted to small forest fragments in one of the most highly threatened portions of the Amazonia (the so-called Arc of Deforestation). This is a typical example of the issues raised by Peloso (2010) and Angulo and Icochea (2010), where lack of taxonomic resolution hinders a proper assessment of threats to cryptic species. For now, we must unfortunately suggest this new species is yet another case of data-deficient taxa—a lot more studies and data are needed to define the conservation status of A. phonotriccus .
REMARKS: Calls of the voucher PV 2597 (specimen no. DT 2123; recordings PLPDR 100; see appendix 3) were unambiguously assigned to A. phonotriccus (“F3” in fig. 6). This voucher was cited as collected in Araguatins (state of Tocantins), on the east margin of Araguaia River. However, this specimen was actually collected in Palestina do Pará, state of Pará—the animal was collected by P.L.V.P. in 2010 (coordinates given by Fouquet et al.’s appendix 1 are correct). The basecamp for this collection site was in Araguatins, and this may have been the source of confusion with the sample provenance (P.L.V.P., personal obs.). Another voucher specimen used in the phylogeny (PV 2412, specimen no. DT 2016), also assigned here to A. phonotriccus (“F3” in fig. 6) was collected in Marabá—this municipality is located in the state of Pará, not Amazonas (AM), as mentioned in appendix 1 of Fouquet et al. (2014).
Of special interest is the acquisition of additional information for the subclade F1, which occurs in partial sympatry with A. phonotriccus (fig. 6). Meanwhile, we conservatively assign only subclade F3 (sensu Fouquet et al., 2014) to A. phonotriccus , and this species is an endemic of the central-east part of the state of Pará in association with the west margin of the lower Araguaia River (fig. 6). With respect to subclade F2, we suspect that it may not be conspecific with A. phonotriccus , given its distribution range associated with another region in the state of Pará (Xingu-Tapajós interfluve; fig. 6). Therefore, we maintain the original status of unconfirmed candidate species of both subclades F1 and F2 for the moment.

| V |
Royal British Columbia Museum - Herbarium |
| T |
Tavera, Department of Geology and Geophysics |
| R |
Departamento de Geologia, Universidad de Chile |
| MK |
National Museum of Kenya |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.