
Malaxis irmae Radins & Salazar, 2014
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.175.3.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03DF465B-FFFE-FFEB-FF4D-FD00CE6BFD7F |
|
treatment provided by |
Felipe |
|
scientific name |
Malaxis irmae Radins & Salazar |
| status |
sp. nov. |
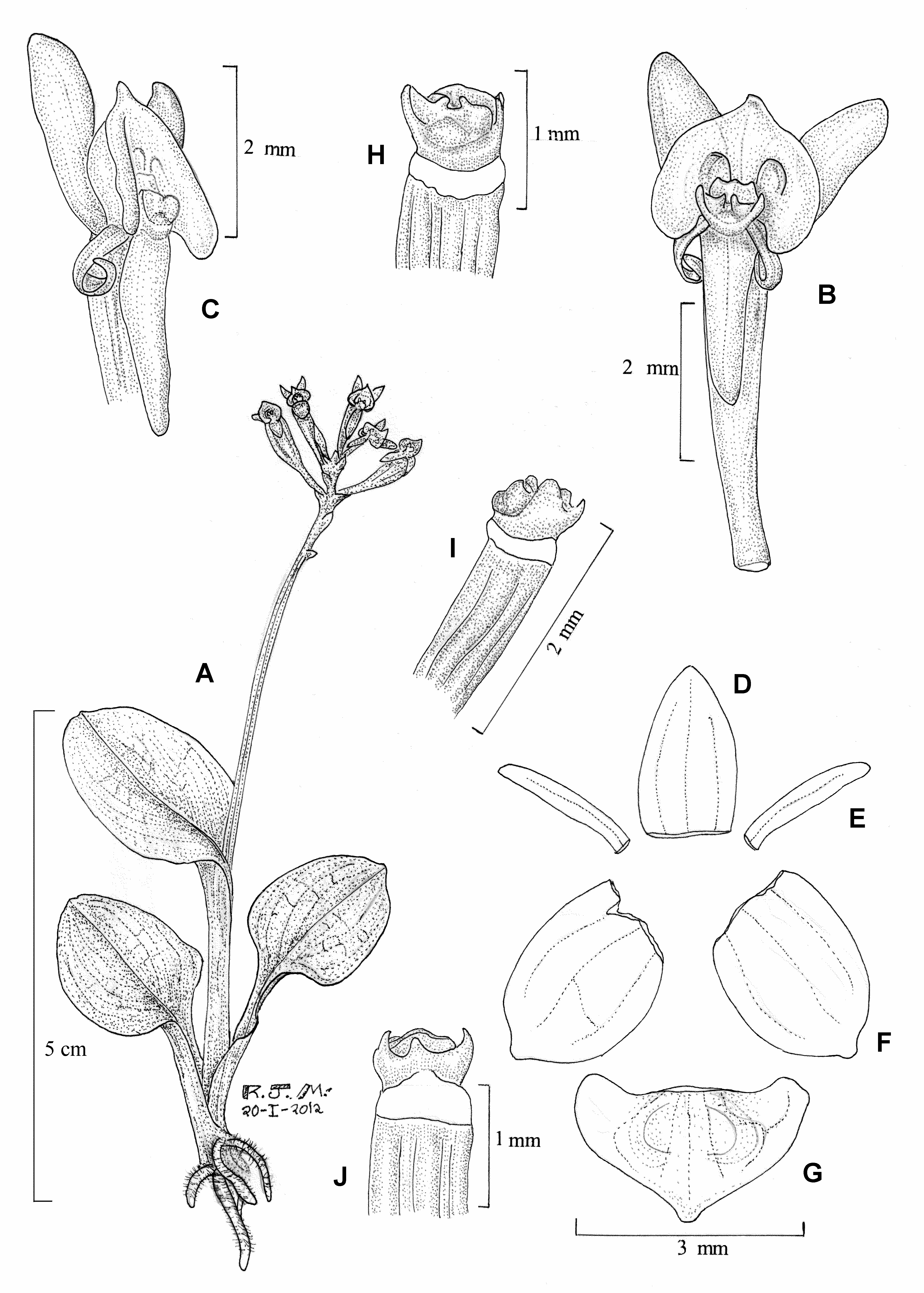
Malaxis irmae Radins & Salazar View in CoL , sp. nov. ( Figs. 3 View FIGURE 3 , 4 View FIGURE 4 ).
Similar to Malaxis cipoensis F.Barros , differing in inhabiting marshy lowland vegetation, 3−5 petiolate leaves per shoot, pale green flowers with somewhat darker green labellum and smaller basal lobules of the labellum.
Holotype:— ARGENTINA. Misiones : Garupá, 11 July 2008, J. A. Radins 105 ( CTES!).
Paludicolous herb 6−14 cm in height including the inflorescence. Roots cylindrical, sparsely pilose, up to 3 cm long, 0.5−1.0 mm in diameter. Rhizome whitish, up to several cm long, 3−5 mm in diameter. Pseudobulbs inconspicuous, ovoid, up to 10 mm long and 6 mm in diameter, when young concealed by the sheathing bases of the petioles. Leaves 3−5 per shoot, ascending, petiolate; petiole white at base but becoming green towards the apex, channelled, distinctly elongate, 12−55 mm long, 3−4 mm wide; blade deep green, broadly ovate to elliptic, base widely cuneate to rounded, apex obtuse-rounded, 14−50 mm long, 10−35 mm wide. Inflorescence provided with a laterally compressed peduncle 50−80 mm long, which is concealed at base by the petiole of the upper leaf; raceme 16−60 mm long, at first condensed and thus appearing somewhat umbellate but the rachis elongates as the flowers open successively and the raceme is cylindrical; rachis slightly angled. Floral bracts slightly concave, incurved, triangular, acute, 1−2 mm long. Flowers non-resupinate; sepals and petals pale green, labellum deep green. Sepals convex, with revolute lateral margins, 3- veined; dorsal sepal adpressed to the ovary, ovate, subacute, 2−2.5 mm long, ca. 1.5 mm wide; lateral sepals diverging, obliquely ovate-elliptic, obtuse, 1.8−2.3 mm long, ca. 1.8 mm wide. Petals strongly recurved, linear, subacute, 1- veined, 2.1−2.2 mm long, ca. 0.2 mm wide. Labellum fleshy, broadly cordate-sagittate, 1.4−1.7 mm long, 1.9−2.8 mm wide; proximal half provided with two rounded excavations; base provided at each side with a slightly retrorse, rounded lobule 0.5−0.6 mm long; apex apiculate, the apicule somewhat incurved in natural position. Column slightly compressed dorsiventrally, ca. 7 mm long and wide; anther dorsal, emarginate; rostellum broadly obtuse; stigma apical, concave, distinctly wider than long. Pollinaria 2, each formed by 2 fused pollinia, ca. 0.5 mm long (fide Izaguirre 2010; not seen). Ovary erect, straight, slightly twisted and somewhat thinner on the proximal one-third, above the middle slightly 6-angled, 6−8 mm long, ca. 1 mm in diameter near the apex. Capsule obovoid-ellipsoid, ca. 6 mm long (plus the pedicel of about the same length), to 5 mm in diameter.
Distribution and ecology:— Malaxis irmae is known only from the Río Paraná and Río Uruguay basins in Uruguay and north-eastern Argentina ( Fig. 5), but it is expected also from southern Paraguay and the state of Rio Grande do Sul, Brazil. It inhabits in water-logged terrain dominated by grasses, and in neighbouring forest edges, between 60 and 100 m above sea level.
Conservation status:— Only three populations of this species have been recorded, but the Argentinian and Uruguayan populations are over 700 km apart in a straight line ( Fig. 5) and it is likely that other populations exist, since suitable habitats (lowland wet grasslands) are widespread over the extensive Río de la Plata basin (which, among others, encompasses the Paraná and Uruguay rivers). At the type locality, urban expansion of the town of Garupá, on the outskirts of the city of Posadas, represents a short-term threat to that population; besides, the wet grasslands and associated water-logged forests that constitute the habitat of this species, are one of the most endangered vegetation types within the Atlantic rain forest biome in south-eastern South America (e.g. Bitetti et al. 2003, Krauczuk 2005, Overbeck et al. 2007). According to the IUCN Red List Categories and Criteria ( IUCN 2012), M. irmae would qualify as Critically Endangered CR (criteria B2a, B2b and C2a(i)). However, both plants and flowers of this species are inconspicuous and can easily go unnoticed, even to trained botanical collectors. Therefore, further field studies are required to attain an objective assessment of its conservation status.
Phenology:— Flowering from March to June. Capsules in advanced stage of development, including some already dehiscing, have been observed from May to August.
Etymology:— The specific epithet honours Ms. Irma Stella Insaurralde, long-term student of the orchids, and the flora in general, of the province of Misiones, Argentina.
Additional specimen examined:— ARGENTINA. Corrientes: Colonia Liebig, D. Boicho s.n. ( CTES!) .
Other records:— URUGUAY. Florida: precise locality not indicated, E. Marchesi s.n. (see Izaguirre 2010) .
Discussion:— Our phylogenetic analysis strongly supports M. irmae as the sister of M. cipoensis ( Fig. 1 View FIGURE 1 , 2 View FIGURE 2 ), in agreement with their similar plant and flower size and overall floral morphology. However, they differ strikingly in habitat preferences, since M. cipoensis inhabits in well-drained soil on rocky field (campo rupestre) areas at 1,000 to 1,340 m elevation, having been found so far only on the Serra do Cipó and Serra da Moeda, both of which form part of the Espinhaço Range in the state of Minas Gerais, Brazil ( Barros 1996, J.A.N. Batista , pers. obs.; Fig. 3F–G View FIGURE 3 ). Malaxis irmae thus differs from M. cipoensis in its lowland, water-logged habitat ( Fig. 4 A–B View FIGURE 4 ), and morphologically it can also be distinguished from the latter by possessing 3−5, distinctly petioled leaves per shoot (vs. 2 shortly petiolate leaves), pale green flowers with a darker green labellum (vs. orangish-green flowers) and smaller basal labellum lobes ( Fig. 4C–E; a View FIGURE 4 good illustration of M. cipoensis for comparison is found in Barros 1996). Malaxis warmingii ( Reichenbach 1881: 64) Kuntze (1891: 673) , a relatively widespread, south-eastern Brazilian species that was not sampled for our molecular analysis, shares with M. irmae a preference for water-logged habitats, the 5−6-leaved shoots and a similar overall floral morphology, but it has much larger plants (40−60 cm in height) and prominent basal labellum lobes ( Fig. 4H–I View FIGURE 4 , Cogniaux 1893 −1896). Malaxis hieronymi ( Cogniaux 1893 −1896: 279−280) Williams (1939: 363) inhabits marshes at high elevations (2,000 −3,500 m) in Bolivia and the Argentinian provinces of Salta, Jujuy and Tucumán, being further distinguished from M. irmae by having pseudobulbs completely covered by fibrous leaf sheaths, two subsessile leaves and labellum lacking basal lobules. The Uruguayan material assigned here to M. irmae was identified by Izaguirre (2010) as M. spicata , a 2-leaved species with prominently lobed, brownish to orange-vermillion labellum provided with a deep triangular cavity, which is limited by a V-shaped, thickened rim. Malaxis spicata is restricted to the Antilles and the southeastern U.S.A. ( Luer 1972).
The overall phylogenetic relationships in Malaxideae are beyond the focus of this paper and will be discussed in detail elsewhere (G. A. Salazar et al., unpublished manuscript). The phylogenetic analysis conducted here was aimed mainly at setting up a context to determine the relationships of the new species, since a morphological comparison with other species of the genus indicated similarities to both M. cipoensis and M. warmingii (see earlier), and its preference for water-logged habitats is shared, besides M. warmingii , by several other species, including South American M. hieronymi (as noted above) and Mexican M. zempoalensis . From our analysis, it is clear that the preference for wet places has evolved more than once in this genus. Although our sample of species of the genus is too sparse to draw conclusions at this time, the relationships recovered suggest that habitat divergence/specialisation may have played a role in promoting speciation in Malaxis s.s., which is exemplified by the contrasting habitat preferences of M. irmae and M. warmingii with respect to M. cipoensis (the closest relative of M. irmae among the taxa we sampled). This and other interesting evolutionary questions, however, will have to be revisited when a more thorough sample of the genus is available for molecular phylogenetic study.
Barros (1996) placed M. cipoensis in Malaxis section Umbellulatae ( Ridley 1888: 315) Barros (1996: 33) and M. warmingii in M. section Spicatae Ridley (1888: 315). The latter is obviously superfluous as it includes the (lecto-) type species of Malaxis ( M. spicata ), whereas our phylogenetic analysis shows that M. cipoensis belongs in the same clade as M. spicata , thus demonstrating that these infrageneric taxa are of little use. These and other infrageneric groups (e.g. those in Margońska et al. 2012), will have to be thoroughly assessed when the phylogenetic relationships in the genus are better understood.



| J |
University of the Witwatersrand |
| A |
Harvard University - Arnold Arboretum |
| CTES |
Instituto de Botánica del Nordeste |
| E |
Royal Botanic Garden Edinburgh |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |