
Phenrica littoralis (Bechyné), Bechyne
|
publication ID |
https://doi.org/10.11646/zootaxa.3686.5.3 |
|
publication LSID |
lsid:zoobank.org:pub:15AEDA97-9392-4A68-A57F-93665D3B8B91 |
|
DOI |
https://doi.org/10.5281/zenodo.6164400 |
|
persistent identifier |
https://treatment.plazi.org/id/03DC8B4E-E005-FFAA-ECBE-A3D7FD4EFD47 |
|
treatment provided by |
Plazi |
|
scientific name |
Phenrica littoralis (Bechyné) |
| status |
|
( Figs1–39 View FIGURE 1 View FIGURES 2 – 7 View FIGURES 8 – 17 View FIGURES 18 – 21 View FIGURES 22 – 27 View FIGURES 28 – 31 View FIGURES 32 – 37 View FIGURES 38 – 39 )
Nephrica littoralis Bechyné 1955: 126 , 1957: 11.
Phenrica littoralis Bechyné 1956: 1018 (new combination), 1958: 660, Scherer 1960: 252; Bechyné & Bechyné 1961: 76.
Diagnosis. Head with impressed punctures on interocular space and close to inner margin of eyes. Elytra piceous with eight yellow spots. Median lobe of aedeagus strongly curved, dorsal median process strongly curved upwards laterally, internal sac with sclerotized plate at base, and larger sclerotized area in the middle. Spermatheca with rounded receptacle, pump smaller, and pronounced neck between both parts.
Redescription. Body oval to elongate, slightly convex ( Fig. 1 View FIGURE 1 ), length 3.57–4.55mm, width 2.60–3.00mm. Color. Head capsule piceous, labrum and mouthparts brown tinged with yellow, apical one-third of mandibles, and maxillary palpi 3–5 dark brown. Antennae piceous, basal half of antennomeres 1 and inner margin of 1–3 flavous. Pronotum yellow. Scutellum, elytra and epipleura piceous; each elytron with four yellow spots, one oval near scutellum, two on the disk, and basal subrectangular one not reaching apical margin. Coxae 1 and trochanter yellowish, coxae 2 and 3, tibiae, and tarsi dark brown, metafemora piceous. Venter, prosternum yellow, mesometasternum piceous, abdomen dark brown tinged laterally with yellow.
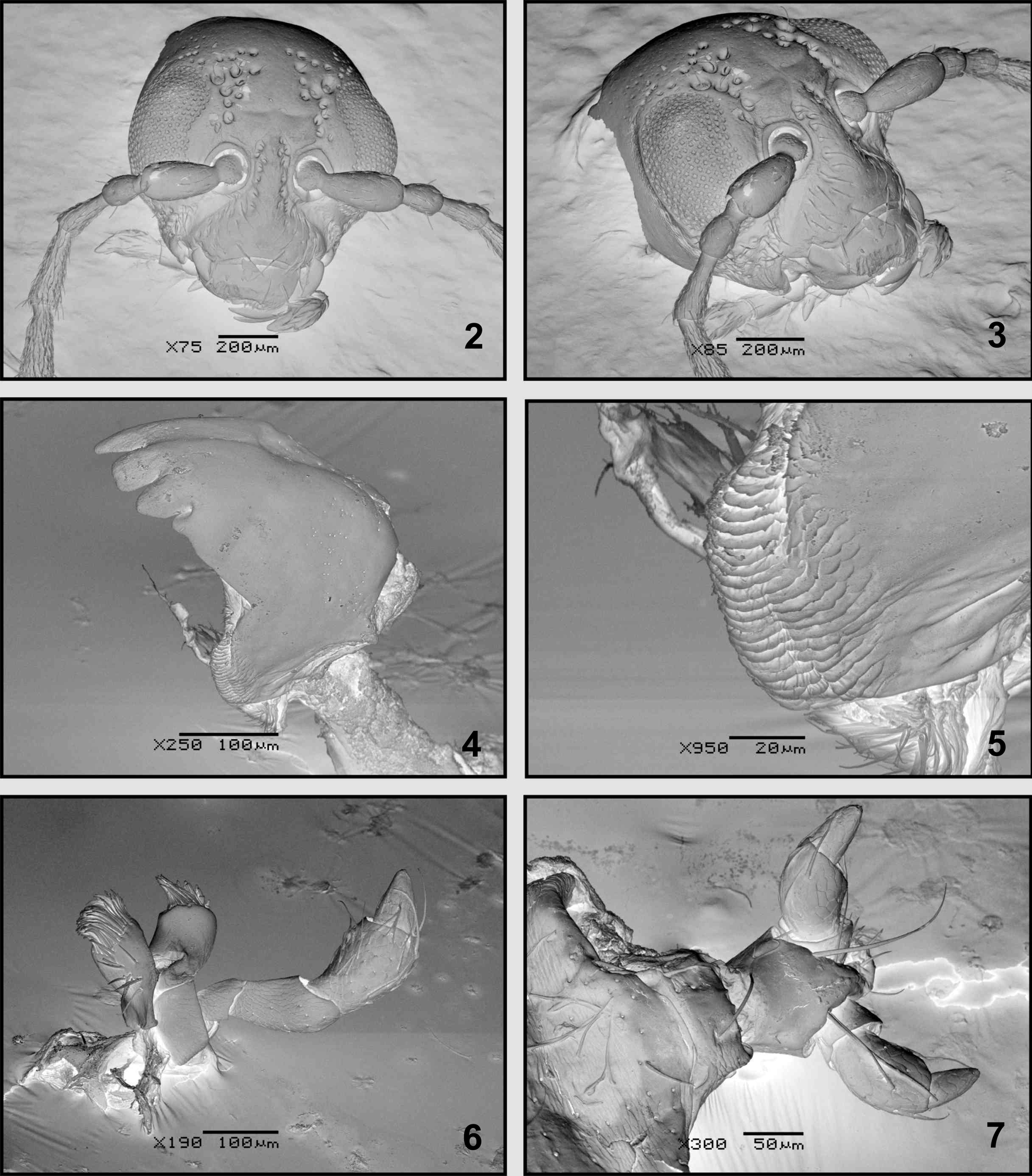
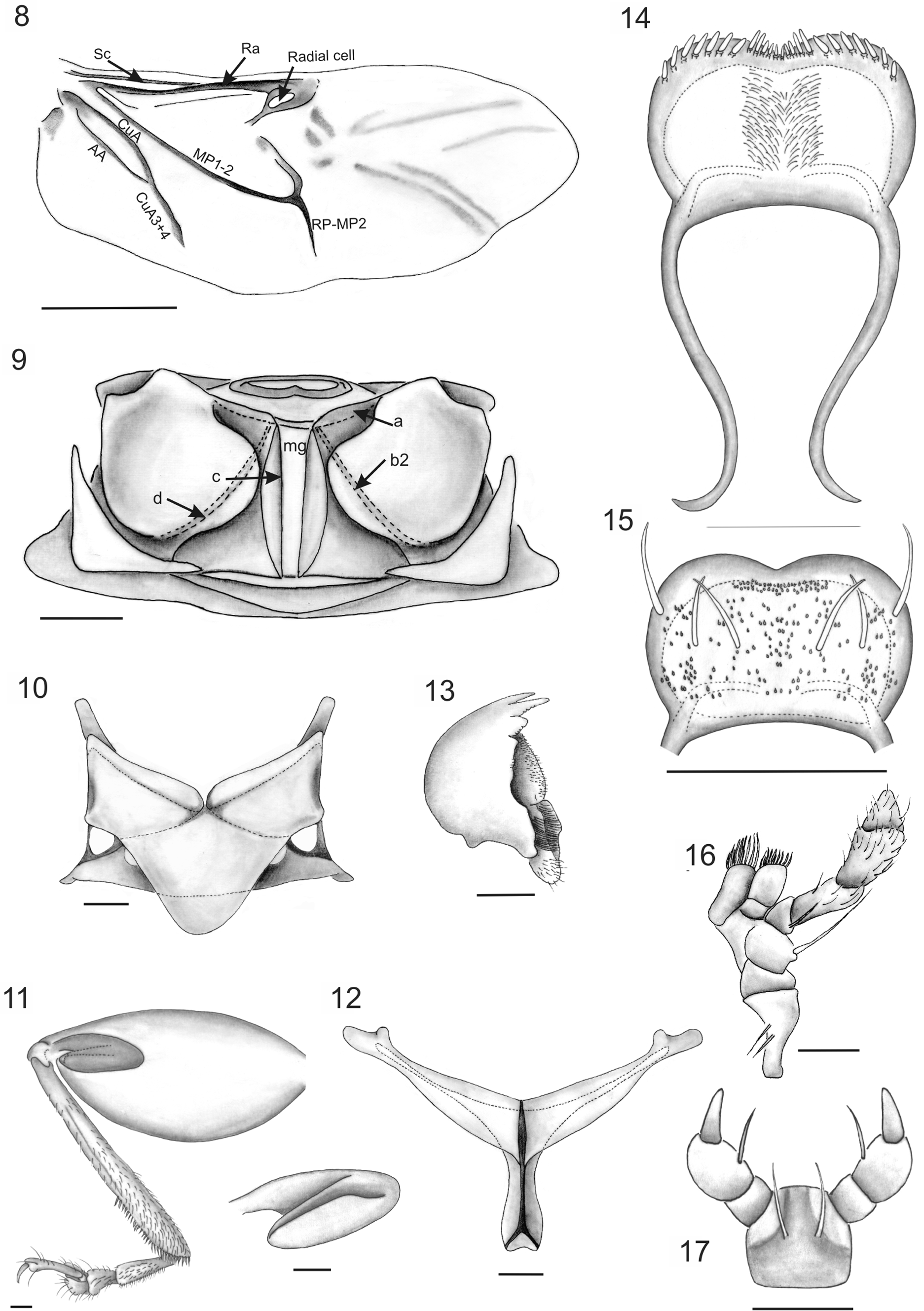
Head ( Figs. 2–3 View FIGURES 2 – 7 ) Vertex smooth; more or less convex in lateral view, with coarse impressed punctures on interocular space near eyes and others close to inner margin of eyes, antennal calli barely raised, trapezoidal, narrower than antennal sockets; supracallinal sulcus and midfrontal sulcus hardly impressed; antennal sockets closed to anterior margin of eyes; interantennal space slightly narrower than transverse diameter of antennal sockets. Eyes convex. Frontal ridge narrow, moderately convex, surrounded laterally by seven long setae, and seven setae below antennal sockets; anterofrontal ridge not separated from frontal ridge, slightly lower than frontal ridge in lateral view. Antennae 11-segmented, inserted below midline of eyes, extending a little more than half length of elytra; antennomere 2 subconical, shorter than 3, antennomeres 4–10 elongate, similar in length, antennomere 11 apically acuminate; antennomeres 1–3 scarcely setose, antennomeres 4–11 densely setose throughout, all antennomeres with erect, sparse setae at apex. Clypeus with 4 preapical long setae. Labrum ( Figs. 14–15 View FIGURES 8 – 17 ) approximately rectangular, lateral margins rounded, anterior margin with small central notch, with a row of six long setae, ventrally eight thick setae in each lateral margin, about eight central, short setae in notch. Mandibles ( Figs. 4 View FIGURES 2 – 7 , 13 View FIGURES 8 – 17 ) 5-toothed, teeth 3–5 visible on external face, teeth 2–3 acute, subequal, tooth 3 more than 2.0 times longer than 4; tooth 4 blunt at apex, almost 2.0 times the length of 5, tooth 5, short, acute, teeth 1–2 visible on internal face, tooth 1 narrow, acute, one-half the length of 2nd, mola ( Fig. 5 View FIGURES 2 – 7 ) well developed. Maxillae ( Figs. 6 View FIGURES 2 – 7 , 16 View FIGURES 8 – 17 ), cardo apically broadened, with two long setae near outer margin; galea and lacinia rectangular, both with fringelike pilosity, scarcer and shorter in the galea. Maxillary palpi with palpomere 1 quadrangular; palpomere 2 short, subquadrate; palpomere 3 subcylindrical, more than one half length of palpomere 2, palpomere 4 subcylindrical, palpomere 5, subconical with wide base, tapering apically, digitiform, subrectangular sensilla patch on externobasal corner. Labium ( Figs. 7 View FIGURES 2 – 7 , 17 View FIGURES 8 – 17 ) with two setae between bases of palps, ligula rectangular. Labial palp with palpomere 1 rectangular; palpomere 2 subconical, more than 2.0 times longer than 1; palpomere 3 subconical with narrow base.
Thorax Pronotum slightly convex, rectangular, PW/PL: 1.61–2.46 times wider than long, widest at middle, PW: 1.70–2.00mm; surface shiny, minutely punctate; anterior margin almost straight, lateral sides slightly expanded anteriorly, posterior margin arched; anterior callosity well produced, rounded, posterior callosity poorly developed, dentiform; each one bearing a long seta. Prosternum convex; intercoxal prosternal process thin, procoxal cavities closed, oval. Scutellum ( Fig. 10 View FIGURES 8 – 17 ). Metanotum transverse, wider than long; metanotal ridge d ( Fig. 9 View FIGURES 8 – 17 ) intersecting c anteriorly to midpoint of c. Metasternum transverse, slightly concave centrally, with slightly pronounced bidentate projection between inner margin of metacoxae; metacoxal cavities inserted at posterior margin, narrowly separated. Metendosternite ( Figs. 12 View FIGURES 8 – 17 , 20 View FIGURES 18 – 21 ) T-shaped, stalk wider at base, shorter than lateral arms, lateral arms widen to apex; mesofurcal-metafurcal tendons well developed, inserted near apex of lateral arms. Hind wings ( Fig. 8 View FIGURES 8 – 17 ) with veins RA, MP, CuA well sclerotized whereas veins CuA2 RP-MP2 and AA scarcely sclerotized. Vein SC connected to RA more than half its length, radial cell darkly pigmented, elongate, subtriangular; RP-MP2 not reaching r4; RP-MP less than one-half the distance of MP 1–2, AA connected to CuA3+4 more than one-half the distance from origin of CuA; cubital anal cell closed, elongate. Elytra oval, convex, surface densely, uniformly punctate, elytra slightly wider than pronotum, HW/PW: 1.16–1.29, humeral calli rounded, slightly prominent, EW/ HW: 1.20–1.29; greatest width near apical one thirds of elytra; epipleura subvertical, basally broad, gradually narrowed at apical half of elytral length; basal inner surface of elytra with an oval binding patch ( Figs. 18–19 View FIGURES 18 – 21 ) uniformly covered with spoonbill-shaped spicules, small sharktooth-shaped spicules covering anterior and posterior end of binding patch, surface close to biding patch covered with thin micro-spicules. All legs similar; pro- and mesotibiae without apical spine, metafemora ( Fig. 11 View FIGURES 8 – 17 ) robust, metatibiae thick, curved, tarsomere 1 of metalegs longer than tarsomeres 2+3 together. Metafemoral spring ( Fig. 11 View FIGURES 8 – 17 ). Tarsal claws ( Fig. 21 View FIGURES 18 – 21 ) appendiculate.
Abdomen Male with apical margin of tergite 7 with about ten long curved setae, base with few scattered microtrichia, indistinct under low magnification; apical margin of sternite 7 ( Fig. 26 View FIGURES 22 – 27 ) weakly truncate, lateral margin with two curved setae. Female with tergite 7 ( Fig. 27 View FIGURES 22 – 27 ) triangular, covered with long setae evenly distributed and six long, curved setae on each lateral margin and 2 rows of 4 curved setae, base densely covered with spiniform microtrichia centrally and compound microtrichia laterally; margin of sternite 7 with four curved setae laterally; and sternite 6 with two lateral, curved setae.
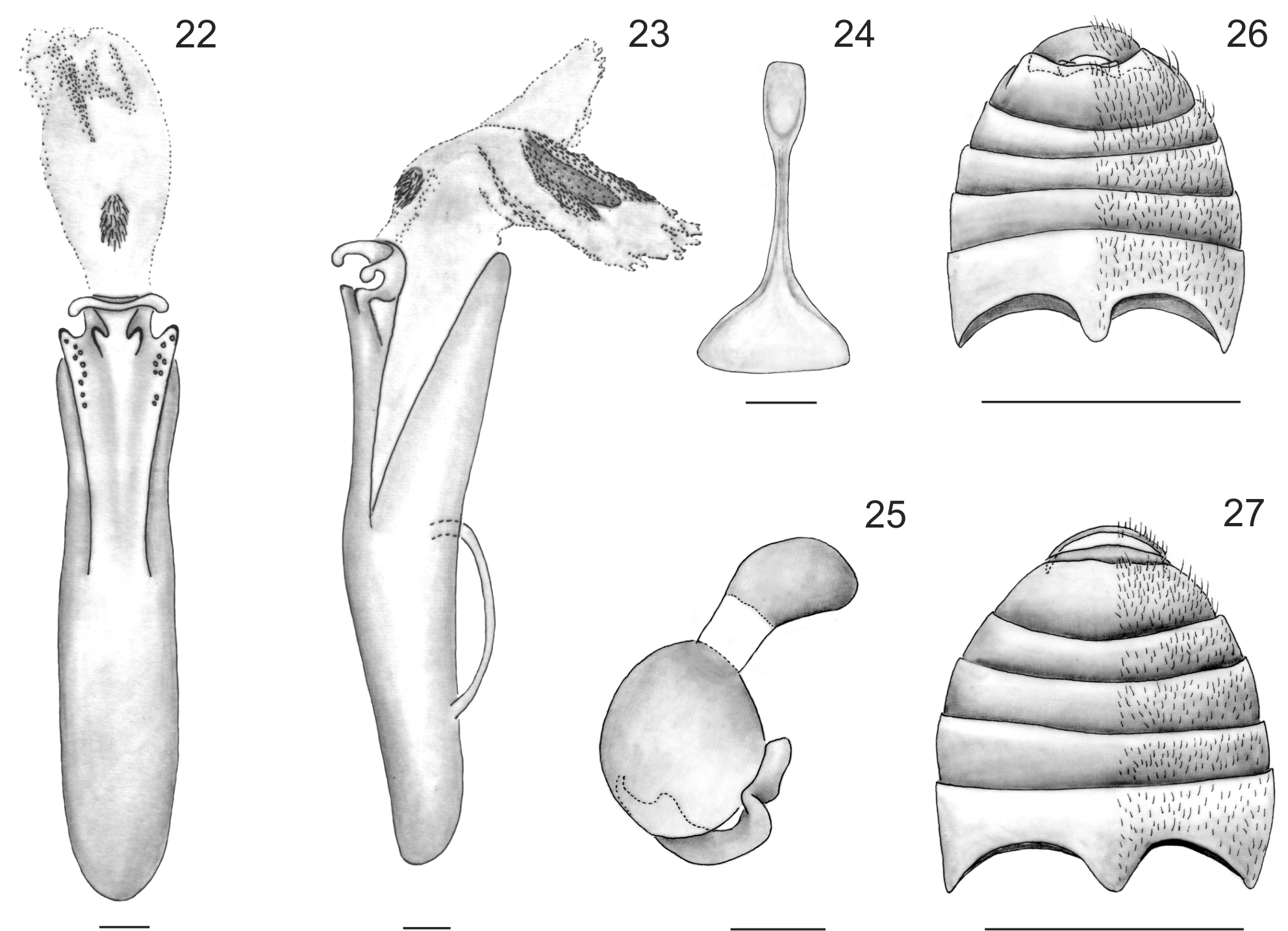
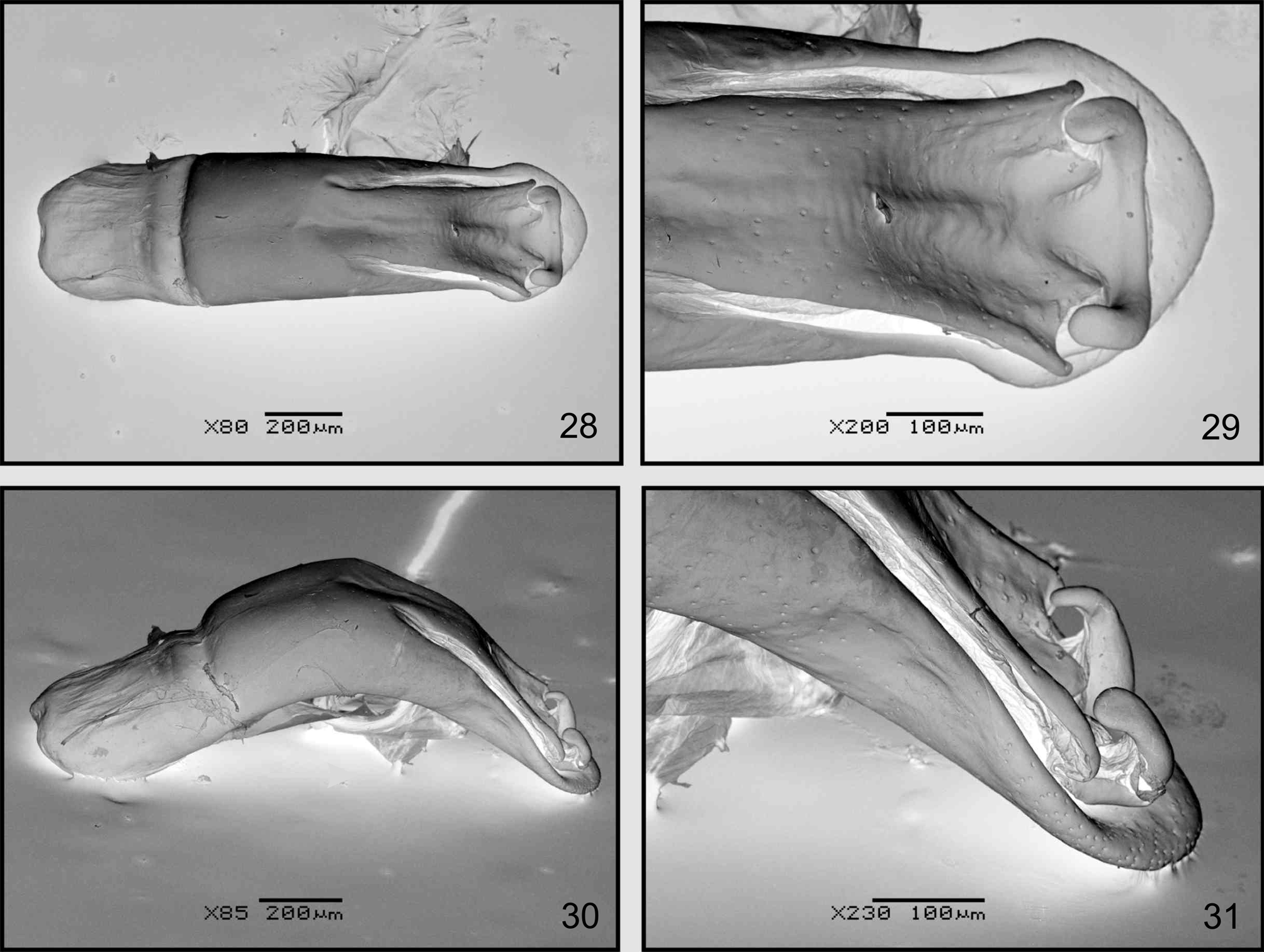
Male genitalia. Median lobe of aedeagus ( Figs. 23 View FIGURES 22 – 27 , 30–31 View FIGURES 28 – 31 ) strongly curved in lateral view. In dorsal view ( Figs. 22 View FIGURES 22 – 27 , 28–29 View FIGURES 28 – 31 ), scarcely wider at apex, ostium relatively large, almost reaching middle of median lobe length, dorsal median process strongly curved upwards laterally; basal foramen wide, internal sac with small rounded sclerotized plate at base and larger sclerotized area in middle of median lobe; tegmen Y-shaped, stem slightly curved, lateral arms longer than stem.
Female genitalia. Sternite 8 ( Fig. 24 View FIGURES 22 – 27 ) with a narrow, basal, rectangular-shaped area, distal area broad, triangular, gonocoxae asetose. Bursa copulatrix membranous, without sclerotized areas. Receptacle ( Fig. 25 View FIGURES 22 – 27 ) and pump of spermatheca well sclerotized, receptacle rounded, pump smaller than receptacle, oval, pronounced neck between both parts, proximal spermathecal duct curved, well sclerotized.
Variability. Specimens from Argentina and northern areas of Brazil (Santa Catarina and Mato Grosso states) are similar in color pattern to the holotype, however, some of these individuals differ by the following traits: presence of a small piceous basal vitta on the thorax, color of abdomen from light brown to yellowish, basal half of metafemora yellowish, tarsi yellowish brown to entirely brown, and elytral spots deeply punctate.
Geographic range. This species was originally described from Brazil (Santa Catarina state), Paraguay and Argentina (Misiones province) ( Bechyné 1955). Later Bechyné recorded this species from other Brazilian states (Bahia, Mato Grosso, Santa Catarina and Rio Grande do Sul ( Bechyné 1956, 1957). The range of Phenrica littoralis includes the Chaco province (Chaco Domain) and Paranense and Cerrado provinces (Amazonian Domain) following the biogeographic scheme proposed by Cabrera and Willink (1980) also denominated Cerrado and Caatinga provinces (Chaqueña subregion) and Bosque Paranaense province (Paranaense subregion) according to Morrone (2001).
Remarks. The original description of P. littoralis in the genus Nephrica included a dichotomous key for six Nephrica species: N. austriaca Schaufuss , N. cisseis Bechyné and N. huebneri Weise (unknown to Bechyné) and the new species: N. aequinoctialiformis , N. weyrauchi y N. mapiriensis . All these species are characterized by the presence of two transverse elytral vittae (each one sometimes divided in two sections) and two rounded spots situated near scutellum. According to the key proposed by Bechyné (1955) and specimens examined from different museums, we conclude that P. littoralis is closely related to P. mapiriensis ( type specimen examined by Cabrera). This species, described from Bolivia, differs from P. littoralis in its longer body, general color dark brown with the ventral surface entirely black, head with only few punctures, prothorax smooth without anterobasal depression, and few isolated punctures next to upper margin of eyes.
Material examined. Type material. Bechyné (1955) described P. littoralis from a female collected in Brazil. Bechyné normally used the word “ Type ” or “ Holotype ” on his identification label to indicate the holotype. This female is labeled as Brasilien, Nova Teutonia, 27º 11º8 52º 23º, Fritz Plaumann typeface/ Type, Nephrica littoralis m., handwriting, J. Bechyné det., 1954 typeface. This specimen is deposited at the Naturhistorisches Museum, Basel, Switzerland ( NHMB).
Additional material examined. ARGENTINA: Misiones: 12 males and 9 females, San Ignacio, 6-XI-03, Sosa and Dorado col. (FUEDEI) ; 8 males and 4 females, Playa del Sol, 23-I-04, Sosa and Dorado col. (FUEDEI) ;
Iguazú, Estación Garganta, 1 male 1 female, Sosa and Dorado col. (FUEDEI) . BRAZIL: Rio Grande do Sul: Serro Azul (actually Cerro Largo), 1 male, VIII-1939, P. Buck col. ( MAPA), 1 male, II-1943, P. Buck col. ( MAPA), 2 males, X-1945, P. Buck col. ( MAPA), 2 females, XI-1949, P. Buck col. ( MAPA), 1 male, Marcelino Ramos, XII-1938, F. Plaumann col. ( MIZA). Santa Catarina: 1 male, Nova Teutonia, II-1935, B. Pohl col. ( MZSP), 2 males and 1 female, V-1935, B. Pohl col. handwritten Phenrica littoralis Bechyné , det. G. Scherer 1954 ( MZSP), 1 female, 12-1939, B. Pohl col., 2˚ handwritten Phenrica littoralis Bechyné , et. B. Pohl 1959 ( MZSP), 3 males, 11- 1944, B. Pohl col. ( MZSP), 1 male, XII-1938, F. Plaumann col. ( MIZA), 1 female, Dirings col, 2˚ handwritten Phenrica littoralis Bechyné , det. Bechyné 1966 ( MZSP), 3males and 4 females, 1975, 27˚ 11’B 52˚ 23’, 300- 500m, Fritz Plaumann, 2˚ handwritten Phenrica littoralis det Scherer 1981 ( DZUP). Mato Grosso: 1 male, Riacho Herval, Rio Paraná, 1-1952, B. Pohl col., handwritten Phenrica littoralis Bechyné , et. B. Pohl 1959 ( MZSP); Itapiranga, 1 female, IX-1953, P. Buck col.( MAPA), 2 females, V-1954, P. Buck col. ( MAPA), 2 males, IX-1954, P. Buck col. ( MAPA), 1 male, X-1954, P. Buck col. ( MAPA), 1 female, I-1960, P. Buck col. ( MAPA); 2 males and 1 female, X-1954 ( MIZA).
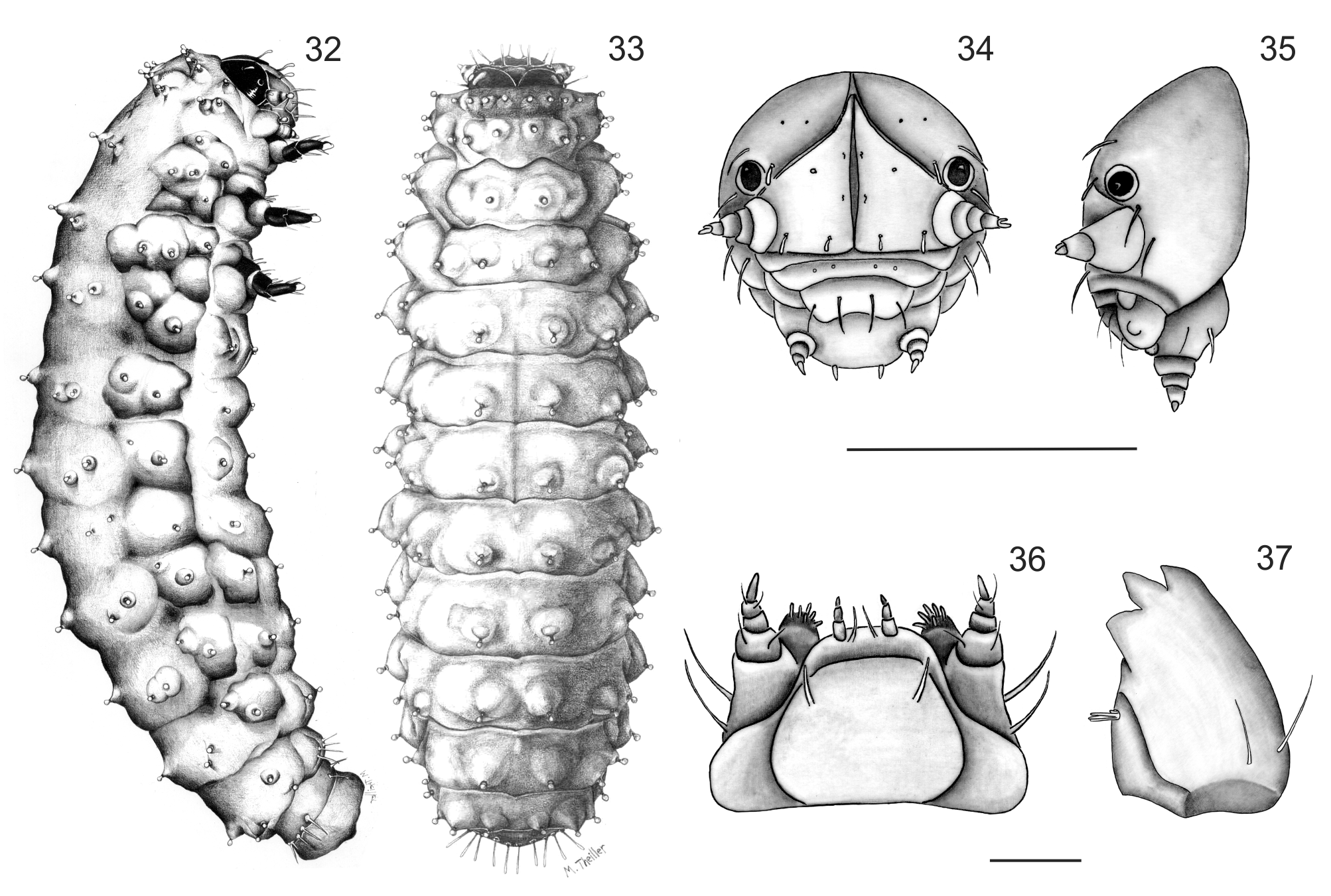
Mature Larvae ( Figs 32–33 View FIGURES 32 – 37 ). Length 3.7–5.6mm, width 1.0– 1.7mm. Body weakly convex when preserved, bright yellow, with prominent scoli each with a seta in dark ring. Head capsule, mouthparts, tubercles, and legs light brown. Microsculpture of the body granulate.
Head ( Figs 34–35 View FIGURES 32 – 37 ). Hypognathous, rounded, well sclerotized. Epicranial suture distinct, Y-shaped; well developed, coronal suture short, extending more than one fourth the length of head, frontal arms pale, distinct throughout, broadly divergent and curved. Endocarina present a black line extending to fronto-clypeal sutures. Frons bearing one pair of long setae medially, two pairs fixed on anterior margin and another two pairs on clypeal area. Each side of epicranial plate bearing two large, dorsal, capitate setae along frontal suture, one fixed laterally on disc, one behind stemmata, and another on the outer side of antennae, four microsetae arranged in a longitudinal row near middle. Antennae short, 3-segmented; attached to head capsule by large, translucent membrane, segment 1 transverse bearing three setae, segment 3 with conical sensory appendage at base. Clypeus transverse, bearing one seta on each lateral margin. Labrum transverse, apical margin emarginated; bearing four large setae in median row. Mandibles ( Fig. 37 View FIGURES 32 – 37 ) robust, heavily sclerotized at apex, 4-toothed, tooth 1 small, teeth 2–3 sharply pointed, tooth 4 blunt at apex, half the length of the previous two teeth, two long mandibular setae inserted dorso laterally, penicillus consisting of four thick setae. Maxillae ( Fig. 36 View FIGURES 32 – 37 ) cardo well-sclerotized, subtriangular, bearing two setae at outer side. Stipes quadrate, with short, narrow inner projection with two long outer setae. Mala densely setose, with about six short setae. Maxillary palpi: palpiger bearing two long setae, segment 1 with two short setae, segment 3 conical. Labium ( Fig. 36 View FIGURES 32 – 37 ) with submentum not much sclerotized, widened at base, with a pair of long, filiform submental setae; prementum broad, with a pair of long setae between labial palpi, labial palpi 3- segmented, short. Hypopharynx densely setose.
Thorax. Pronotum transverse, pronotal sclerite not well defined, dorsal side with short club-like setae each borne on a minute tubercle arranged in anterior and posterior rows, epipleural tubercle bearing three setae, small pre-hypopleural tubercle bearing one seta. Meso- and metathorax wider than prothorax, each bearing four setae arranged along mid-line. Meso-and metapleura each with alar tubercle bearing two setae and anterior epipleural bearing one seta, pro-meso-and metasterna with two medial setae. Spiracle annuliform displaced into mesothoracic region. Legs 5-segmented, strongly chitinized, the third pair slightly increased in size; coxa trapezoidal bearing 3 long setae and a sensilla; trochanter, triangular, with two setae, femur sub-rectangular bearing eight long setae; tibia bearing six setae. Tarsungulus moderately curved, with single basal seta; paranonchial appendix projecting behind tarsungulus.
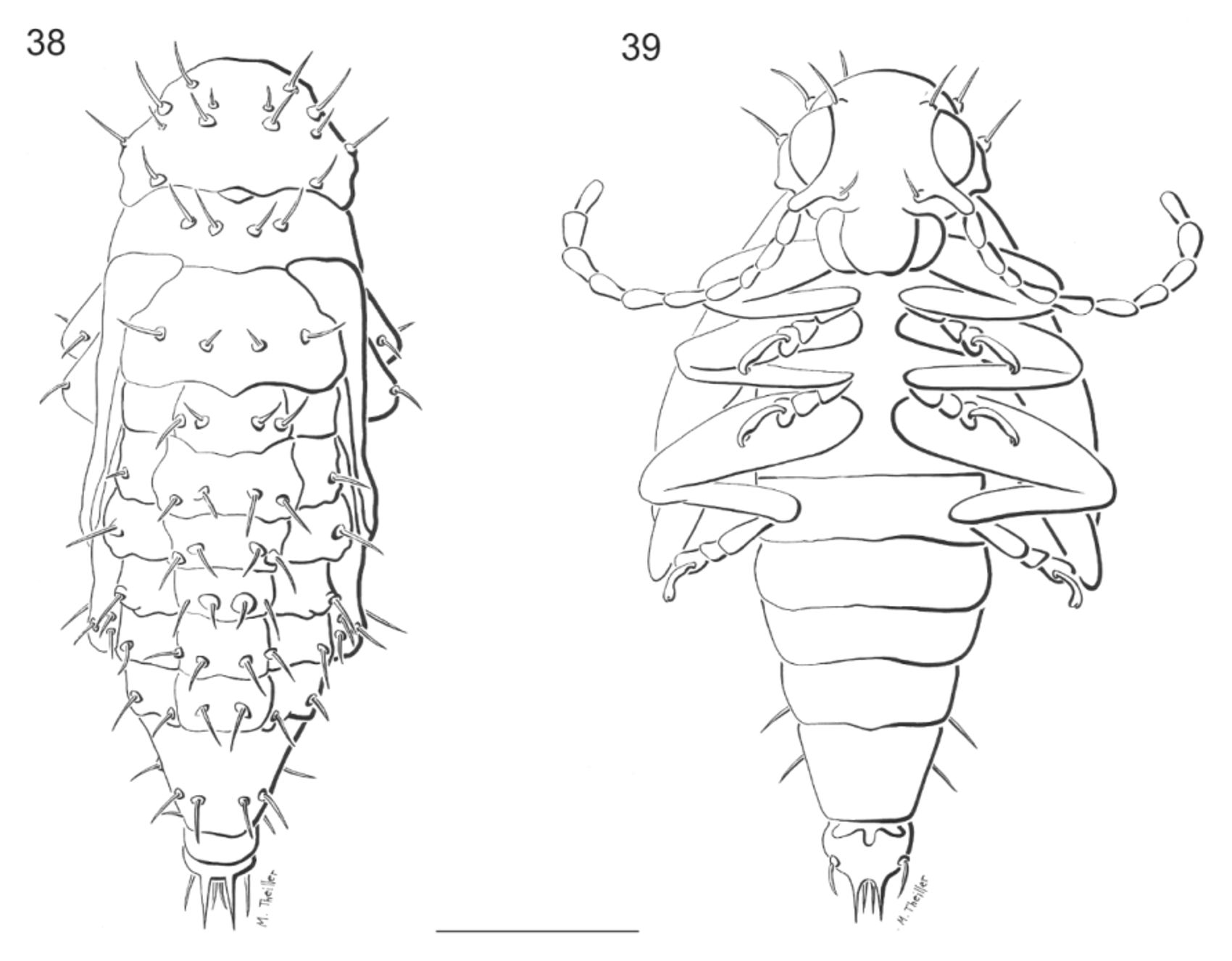
Abdomen. Abdominal segments 1–8 with a pair of dorsal setae, posterior parascutal area bearing two setae, epipleural and hypopleural areas each bearing one seta, ventrally eusternal area of segments 1–7 with a pair of very short club like setae; segment 8 with four pairs of long, filiform setae segment 9 forming fleshy pygopod. Spiracles 1–8 annuliform.
Pupa ( Figs. 38– 39 View FIGURES 38 – 39 ). Length 3.4–4.1mm, width 1.8–2.5mm. Color of body pale yellow. Tubercles, base of spiracles and cerci light brown. Head deflexed, not visible from above, with only two pairs of setae: one on eyes and one subantennal. Mouthparts well developed. Pronotum with seven pairs of pronotal setae situated on disc and margins: two pairs on disc, three pairs on outer margins, and one pair on posterior margin, one pair of rounded spiracles. Meso- and metathorax with two pairs of meso-and metathoracic setae arranged in transverse rows. Podothecae with two pairs of femoral setae on apex of femora. Abdominal segments1–6 transverse, each with two pairs of dorsal setae one pair of spiracles and one pair of pleural setae originating from a lateral projection, abdominal segment 6–7 with similar chaetotaxy but without spiracles, abdominal segment 7 subconical. Abdominal segment 8 reduced with two short lateral setae, abdominal 9 with one pair of urogomphi straight with two pairs of setae at base, ventrally two pairs of lateral setae and other two shorter setae medially.
Remarks. Considering all known larvae of Disonychina , Phenrica is more closely related to those of Disonycha . Lawson (1991) illustrated larvae of Disonycha xanthomelas (Dalman) , D. triangularis (Say) and D. alternata Illiger , however the lack of detailed chaetotaxy characters for these descriptions prevent in-depth comparisons with P. littoralis . Phenrica littoralis shares the following characters with these disonychine species: similar arrangement of body setae, head and legs brown, antennae 2-segmented, stemmata present, abdomen with prominent light scoli. Larvae of P. littoralis differs by the shape of mandibles, maxillae, tarsunguli and the number of body setae.
Little is known about Disonychina pupae. The pupae of only five species of Disonycha have been described by different authors ( Chittenden, 1899; Frers, 1919; Hemenway & Whitcomb, 1968; Whitehead, 1918). However, only the descriptions of D. argentinensis Jacoby , D. xanthomelaena and illustrations of D. bicarinata Boheman (the three latter modified by Cox 1996, 1998) can be compared with Phenrica . Disonycha glabrata (Fabricius) has not been described in detail and is therefore impossible to properly compare.
The pupa of P. littoralis differs mainly from D. argentinensis by the presence of seven pairs of setae on the pronotum, femora bearing two setae, abdominal segments with two pairs of dorsal setae and one pair of pleural setae, ventral surface without setae and urogomphi straight. It is difficult to compare in detail with the pupa of D. xanthomelaena , but according to the characters presented by Cox (1996), differences were observed by the number of setae on the pronotum, abdominal sternites, and the urogomphi slightly divergent. According to the illustrations of D. bicarinata , this species shares with Phenrica the number of setae on the pronotum, abdomen and femora.
Phenrica . littoralis differs by its more slender body, distribution of head setae, the conical shape of abdominal segment 7, and number of its setae and urogomphi straight and close together.
On the other hand, some pupal features, like the position and presence of some setae and urogomphi, allow conjecturing as to the pupation biology of this species ( Cox 1996). According to this author, the chief function of both structures is to protect the pupal integument from abrasion against the walls of the cell. Therefore, it should be present in species that build cells. Laboratory observations indicate P. littoralis pupates in the soil substrate in a cell probably built with soil cemented with some excretion (n=23). It was also observed that some individuals pupated even if deprived of a substrate (sand or soil), but emergence success was much lower (2/10). These observations are similar to those for Disonycha spp., which pupate in wet soil and build a mud cell sealed with cement-like anal or oral exudates, or both, applied in successive layers ( Vogt et al. 1979).
Host specificity. None of the Phenrica species are listed as pests in Argentina ( Cabrera 2004) or Brazil ( DÁraujo e Silva et al. 1968), but some species recorded on Amaranthaceae ( Irene , Cyathula and Althernanthera) ( Jolivet & Hawkeswood 1995), were also observed on cultivated Fabaceae on which they occasionally caused some damage ( Bechyné & Bechyné 1966). Phenrica guerini (Bechyné) was the first species in the genus mentioned as a biocontrol agent of another weed, Barbados gooseberry, Pereskia aculeata Miller (Cactaceae) ( Klein 1989).
Biological aspects. Phenrica littoralis was found in San Ignacio, Misiones and in several locations nearby. In the field, adults were found in large numbers on shaded plants growing under trees in a completely terrestrial situation. Larvae have not been observed in the field. In the laboratory, egg masses were obtained on moist tissue paper (14.5 ± 5.7 eggs/mass, n=12). Around 85% emergence occurred between 9 and 12 days after oviposition. Eggs are yellow and larvae and pupae are light pink. Larvae fed on alligator weed leaves and pupated in the substrate. Immature stages lasted between 24 and 35 days (28.3 ± 4.3 days, n=23), and the larval survival was 46 % (23/50). Immature duration and biological aspects are similar to its close related species P. g u e r i n i ( Klein 1989).
Final considerations. Duckett (1999) proposed the first phylogenetic hypothesis of relationships among the subtribe Disonychina based on adult morphology. In this analysis it is postulated that species of Balzanica , Disonycha and Phenrica constitute a clade supported mainly by genitalic characters. Duckett (1999) further proposed that the series Disonycha should include the genera Phenrica and Disonycha and that Phenrica may be a junior synonym of Disonycha . Although as only one species of Phenrica was included, a detailed analysis of the genera is still needed.
This is the first description of the immature stages of Phenrica , which will contribute to enhance the taxonomic and biological knowledge of Alticini . As a complement of the study of adult structures, the morphology of immature stages provide a source of new characters that could help establish a phylogenetic hypothesis of relationships among taxa.
Other flea beetles were proposed as potential candidates for the biological control of Alternanthera philoxeroides ( Julien & Chan 1992; Sosa et al. 2004), but these species were not sufficiently specific. Phenrica littoralis was only found on terrestrial alligator weed and preliminary information from laboratory (unpublished data) encourages new studies. The beetle has being introduced into quarantine facilities in Australia to study its host specificity.
Agasicles hygrophila Selman & Vogt is the first flea beetle used as biological control agent of alligator weed. If a high-level of specificity of P. littoralis is confirmed it will be the second flea beetle – utilized for the biological control of Alternanthera philoxeroides worldwide.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Phenrica littoralis (Bechyné)
| Cabrera, Nora, Sosa, Alejandro J. & Julien, Mic 2013 |
Phenrica littoralis Bechyné 1956: 1018
| Bechyne 1961: 76 |
| Scherer 1960: 252 |
| Bechyne 1956: 1018 |
Nephrica littoralis Bechyné 1955: 126
| Bechyne 1955: 126 |