
Hauserimyia, Stuke, 2019
|
publication ID |
https://doi.org/10.21248/contrib.entomol.69.2.301-317 |
|
DOI |
https://doi.org/10.5281/zenodo.3716552 |
|
persistent identifier |
https://treatment.plazi.org/id/03DB87D7-6E12-3903-FF27-FE6AFB40F945 |
|
treatment provided by |
Plazi |
|
scientific name |
Hauserimyia |
| status |
gen. nov. |
Hauserimyia View in CoL gen. nov.
( Figs 14−27 View Figs 14–16 View Figs 17–20 View Figs 21–24 View Figs 25–27 )
Type-species: Hauserimyia martini View in CoL spec. nov., designated herewith.
Diagnosis: Hauserimyia is easily identified as belonging in the Conopinae because of the characteristic shape of the antenna, which has an apical stylus. Given the combination of an elongated pedicel ( Fig. 15 View Figs 14–16 ), an elongated basal flagellomere ( Fig. 24 View Figs 21–24 ), a lack of ocelli, a very short (i.e. distinctly shorter than head length) and apically widened proboscis and more than 10 lamellae on the labrum ( Fig. 15 View Figs 14–16 ), there are only three other similiar genera which occur in the Oriental and Palaearctic regions. These are Leopoldius RONDANI, 1843 , Abrachyglossum KRÖBER, 1919 and Neobrachyglossum KRÖBER, 1915 . Hauserimyia is easily distinguished both from these and from all other conopid genera by the unique shape of the arista, which has three large aristomeres as shown in Fig. 24 View Figs 21–24 , and the unique pairs of warty submedial protuberances on tergites 3 and 4 of the female ( Figs 17−19 View Figs 17–20 ).
Etymology: The genus Hauserimyia is derived from the name „Hauser“ in combination with the Greek noun „myia“, meaning “fly”. This patronym is dedicated to Martin Hauser (Sacramento) to whom I am very grateful for his long-standing support of my dipterological work. The name Hauserimyia is to be treated as feminine.
Description: Head: Arista stylus-like, with 3 aristomeres situated at tip of basal flagellomere ( Figs 15 View Figs 14–16 , 24). Basal aristomere and medial aristomere short and about the same size, apical aristomere elongated, tapering gently to a blunt tip. Scape about 1.5 times longer than maximum width, dorsally with black setae. Pedicel about six times longer than maximum width, completely covered with long black setae, with a small and inconspicuous blunt ridge at base, and slightly expanded towards apex. Basal flagellomere long, about three times longer than high and almost as long as pedicel. Basal flagellomere pointed, ventrally lacking a membranous area, and lacking setae dorsally. Lunule between base of antennae and ptilinal suture distinct, slightly shorter than width of scape. Eye brown, lacking ommatrichia. Facets all of about the same size. Posterior margin of eye convex, lacking an indentation. Gena height / eye height (measurements are taken from head in lateral view) = 0.2. Ocellar tubercle not evident. No ocelli and no ocellar triangle evident. Black frontofacial spots distinct. Vertex as broad as frons, not separated from latter by a ridge, distinctly rounded anteriorly, and covered with black setulae on posterior only. Gena with a few black setulae, facial ridge lacking setulae. Distinct facial grooves reaching mouth edge. Distinct facial carina reaching from base of antennae to a distinctly widened and outstanding frontoclypeal tubercle. Ptilinal suture stretching a short way on either side beneath the antennal bases. The area surrounding the ptilinal suture is yellow with the exception of the lunule. Mouth opening tapering dorsally. Postcranium not obviously invaginated. Postgena not widened and therefore not separated from the occiput. Bottom portion of postcranium distinctly separated. Palp present, but very small and hard to detect in dried specimens. Proboscis geniculated once only, at base. Labium shorter than head-length, hardly projecting out of mouth opening, distinctly thickened basally, the anterior section not fused into a tube. Labrum only evident in about the basal half of the haustellum. Labellum short, completely divided, broad (at least two times broader than the adjacent haustellum) and covered with long yellow setulae. Labellum with about 15 distinct pseudotracheae on each side. Head lacking setae.
Thorax: Presternum distinct, slightly wider than the compound basisterna. Presternum distinctly separated from basisterna. Basisternum broad, narrowed to a tip, lacking setulae. Proepisternum with a hardly visible small black seta which is adpressed and directed posteriorly, and which could not be found in every specimen. Mediotergite convex, lacking setulae, and hardly projecting over scutellum. Subscutellum inconspicuous. Metakatepisternum, anepisternum and anepimeron lacking setulae. Subcostal-radial crossvein sc-r present ( Fig. 21 View Figs 21–24 ). Radial-medial crossvein r-m complete. Radius R 1 and R 2 terminate close together in costa, well beyond end of subcosta. Radius R 4+5 with a shallow and even curve in the distal section that is directed towards the wing tip. Radial cell r 4+5 pedunculate, with vein R 4+5 + M distinctly expressed and about as long as radial-medial crossvein r-m. Cubital cell cup elongated (distinctly longer than vein A 1 +CuA 2) and pointed distally (cubitus CuA 2 and anal vein A 1 meet at an acute angle). Cubital vein CuA 1 and crossvein bm-cu distinctly separated. Alula broad (about 2.5 times broader than long), with brown setulae on posterior margin. Venae spuriae pronounced in radial cell r 4+5 and also indistinctly in cubital cells cup and cua 1. Base and stem of haltere each with areas of sensillae. Knob of haltere with isolated black setulae. Posterior surfaces of fore and middle tibiae lacking obvious dusted fields distally. Areas with dense brown setulae ventrally at tip of fore tibia and ventrally and posteriorly at tip of hind tibia. Middle femur posteriorly lacking regularly arranged long setulae. Hind femur dorsally lacking outstanding long setulae. No preapical setulae dorsally on tibiae. No setulae ventrally on tibiae. All femora ventrally with two rows of short, regularly arranged black setae. Hind femur not obviously thickened in basal half. Line of black, regularly arranged setae posteroventrally on fore metatarsus, anteroventrally and posteroventrally on middle metatarsus, and anteroventrally on hind metatarsus.
Abdomen: Tergites 1−3 fused but distinctly separated from each other ( Figs 17, 18 View Figs 17–20 ). Lateral margins of tergites almost straight.
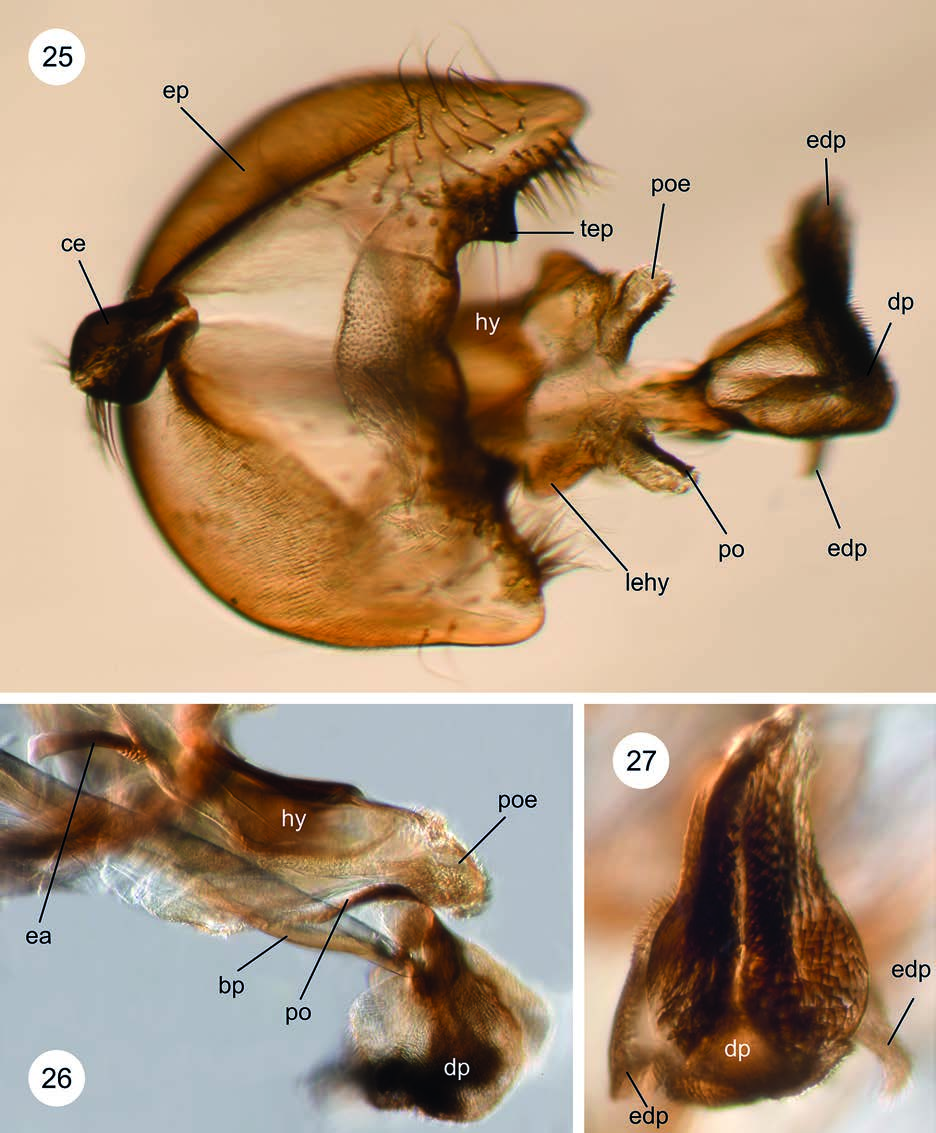
♂ preabdomen with 5 tergites. Tergite 1 with obvious black setulae laterally on the bulbous projections. Tergite 2 hardly elongated but parallel-sided, about 1.5 times as long as broad when viewed dorsally, and lacking obvious lateral tufts of setulae. Tergite 3 slightly widened posteriorly, about 1.5 times as wide posteriorly than anteriorly, and not obviously broader than long.Sternites 1−5 present, sternites 1 and 2 not fused and distinctly separated. Tergite 5 and sternite 5 distinctly separated. Sternite 5 apically with a small field of thick setulae. Protandrium obviously broader than epandrium and therefore projecting over it. Sternite 8 distinctly delimited from protandrium. Ventrally the lateral edges of the protandrium are fused by a narrow sclerotised strip, which is not widened medially. Paired cerci distinct, completely sclerotised and covered with setulae ( Fig. 25 View Figs 25–27 ). Posterior and anterior surstyli absent. No obvious strong black setulae nor long black setulae which would indicate the base of a surstylus. Subepandrial plate not sclerotised and therefore not evident, covered with microtrichia. Dorsal hypandrial bridge developed. No hypandrial lobe evident. Hypandrial bars fused distally and hypandrium ending in a hypandrial arm. Hypandrial membrane reduced, lacking microtrichia. Phallus sheath fused dorsally; apically with lateral evagination ( Fig. 25 View Figs 25–27 : lehy) close to the fused postgonite evagination, which has several short setulae. Postgonite distinct. Postgonite evagination hardly sclerotised, not projecting above distiphallus, and bearing distinct microtrichia ( Fig. 26 View Figs 25–27 : poe). No plate on inner side of postgonite evagination but dense elongated and lightly sclerotised setulae. Ring sclerite developed. No epiphallus recognised. Phallus apodeme longer than hypandrium arm. Ejaculatory apodeme elongate, with a distinct narrow attachment to sperm sac.
♀ abdomen with sternites 1−2 fused. Sternites 3−4 not protruding ventrally, posterior parts of sternites inconspicuous. Tergites 3 and 4 each with a conspicuous pair of submedial protuberances, the surfaces of which bear some wart-like structures ( Figs 17−19 View Figs 17–20 ). Tergite 5 and sternite 5 fused laterally to form a syntergosternite with an obvious theca below ( Figs 20 View Figs 17–20 , 22 View Figs 21–24 ). Posterior part of sternite 6 inconspicuous, slightly bent dorsally ( Fig. 23 View Figs 21–24 ). Tergite 7 distinctly bent ventrally, without a longitudinal gap and lacking a protruding tooth in middle of posterior margin. Sternite 8 not fused with syntergite 8+9 and therefore not connecting on its sides. Tooth on syntergite 8+9 distinct, its base elongated anteriorly. Sternite 9 bulging posteriorly, covered with strong long black setulae, and with long setulae on posterior margin. Paired cerci distinct. Sack-like ventral protrusion of vagina with distinct annular sclerotisation. Sack-like ventral protrusion is hardly larger than annular sclerotisation and no evagination or additional sclerotisations are recognised. 2 pairs of round spermathecae, spermathecal ducts fused shortly after leaving spermathecae. Spermathecal ducts only sclerotised directly at spermathecae.
Distribution: The genus Hauserimyia is known only from the locus typicus of Hauserimyia martini in the Min Mountains (Minshan). The biogeographical classification of the locus typicus is discussed under Gellergrimmellus , above.
Biology: Nothing is known about the biology of this genus.
Phylogenetic placement of Hauserimyia : Without doubt Hauserimyia belongs in the Conopinae due to the following characters: stylate arista placed apically on basal flagellomere ( Fig. 15 View Figs 14–16 ); ocelli and ocellar tubercle absent; vertex reaching from one eye to the other ( Fig. 16 View Figs 14–16 ); no ocellar triangle; lunule distinct ( Fig. 15 View Figs 14–16 ); facial grooves reaching to mouth opening, divided by a central carina which widens ventrally; mouth opening tapering dorsally; postgena not widened and therefore not separate from the narrow occiput; chaetotaxy reduced, no obvious setae on the head; no preapical setulae dorsally on the tibiae; radial cell r 4+5 petiolate and R 4+5 + M distinct ( Fig. 21 View Figs 21–24 ); subcosta-radial crossvein sc-r well developed ( Fig. 21 View Figs 21–24 ); ♂ protandrium obviously broader than epandrium and therefore projecting over it ( Fig. 14 View Figs 14–16 ); ♂ hypandrium sheath fused dorsally and therefore surrounding the phallus completely.
The further subdivision of the Conopinae is not currently resolved. GIBSON & SKEVINGTON (2013) attempted to separate the Conopinae into eleven tribes based on a phylogenetic approach but this took into account only relatively few species, and did not sufficiently discuss the morphological characters used in the cladistic analysis ( BORKENT 2018). It is therefore more of a preliminary basis for discussion than an elaborated hypothesis concerning the evolution of the Conopinae . STUKE (2017) does not recognise any tribes in the world catalogue of Conopinae and it has been demonstrated that the system proposed by GIBSON & SKEVINGTON (2013) cannot be applied consistently in several cases ( STUKE 2014, 2018). It is therefore not currently possible to place Hauserimyia phylogenetically with any sister taxon within the Conopinae in the absence of a complete cladistic analysis of the subfamily. Nevertheless the similarity between Leopoldius , Neobrachyglossum and Hauserimyia is striking and it may well be the case that these form a monophyletic group.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.