
Metallactus corruptus Suffrian, 1866
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4413.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:D65DAEB6-0499-44F9-A70D-0720BA71D520 |
|
DOI |
https://doi.org/10.5281/zenodo.5970566 |
|
persistent identifier |
https://treatment.plazi.org/id/03DB8796-3219-9805-FF48-1190FD73FE51 |
|
treatment provided by |
Plazi |
|
scientific name |
Metallactus corruptus Suffrian, 1866 |
| status |
|
Metallactus corruptus Suffrian, 1866
( Figs 6 View FIGURES 5–6 ; 25 View FIGURES 20–28 )
Metallactus corruptus Suffrian, 1866: 370 ; Clavareau, 1913: 92; Blackwelder, 1946: 641.
Type material. The following specimens which fit in well with the data in the original description were tracked down: 1♂ 4♀♀ in MNHUB; 1♂ 1♀ in MLUH; 5♂ 4♀♀ in NMV; 1♂ BMNH. The typification has been made as follows. I would have liked to designate as lectotype a specimen in NMV, which possess the greatest number of syntypes. As a matter of fact, it is quite sure that the material in NMV belongs to the assemblage studied by Suffrian in his 1866 monography, because the only species of Metallactus housed in NMV are the ones which Suffrian pointed out to be housed in NMV. Therefore, it does not seem that since then other studied and determined material has been added to NMV general collection. However, none of Vienna's specimens brings clues of original labels, because all of them have been replaced by rather recent printed labels, certainly attached much after Suffrian's study. In the specific case of Metallactus corruptus , all of the specimens carry the indication: // “Trisanga Brasilien Natterer” //. However, it should be noted that “Trisanga” is certainly a misspelling for Irisanga (= Orissanga), in the State of São Paulo, a site undoubtedly visited by Natterer ( Schifter, 1993). In the original description, Suffrian does not mention specific localities nor the name of the collector, as generally did in other cases. In the light of the foregoing considerations, I am convinced that Suffrian, on this occasion and in other similar cases (see below), didn't fully report all the collection data, or some information was unavailable to him when he got the material in study, nonetheless the non-correspondence between the text and the label collection data does not allow a formal and undeniable designation of the material in question as belonging to the type series of the species. Unfortunately, this is a recurring problem in detecting properly the type series of some species described by Suffrian. For all these reasons the specimen housed in BMNH, whose data collection match better the original description, was designated as the LECTOTYPE: ♂, glued, detached abdomen on the same label, aedeagus on a separate label, // [small white label handwritten unreadable] // “55 23” [round white label, handwritten] // “Type Suffr Coll Baly” [white label, handwritten] // “Baly Coll.” [white label, printed] // “ Metallactus corruptus Suffr Brazil ” // “ Metallactus corruptus Suffrian, 1866 LECTOTYPUS D. Sassi des.” [red label, printed] //. PARALECTOTYPES are as follows. 5♂ 4♀♀, // “Trisanga Brasilien Natterer” [white label, printed] ( NMV); 1♂ 1♀, // “corruptus m. Brasil ” [blue label, handwritten] // “28012” and “28013 [white label, handwritten] // “MLU Halle WB Zoologie S.Nr. 71116” [white label, partly printed] ( MLUH); 1♂ 4♀♀, // “ corruptus Suffr. * ♂ ♀ Brasil Virm.” [blue label, handwritten] // “Hist.-Coll ( Coleoptera ) Nr. 23985 Metallactus corruptus Suffrian Brasil., Virmnd. Zool. Mus. Berlin” [green label, printed] //. The number of specimens and the label information match the registration data from the old catalogue of the MNHUB (“23985 corruptus Suffr. 5 Brasil Virmnd”).( MNHUB). All paralectotypes were labelled: // “ Metallactus corruptus Suffrian, 1866 PARALECTOTYPUS D. Sassi des.”[red label, printed] //.
Type locality. “ Brazil ”
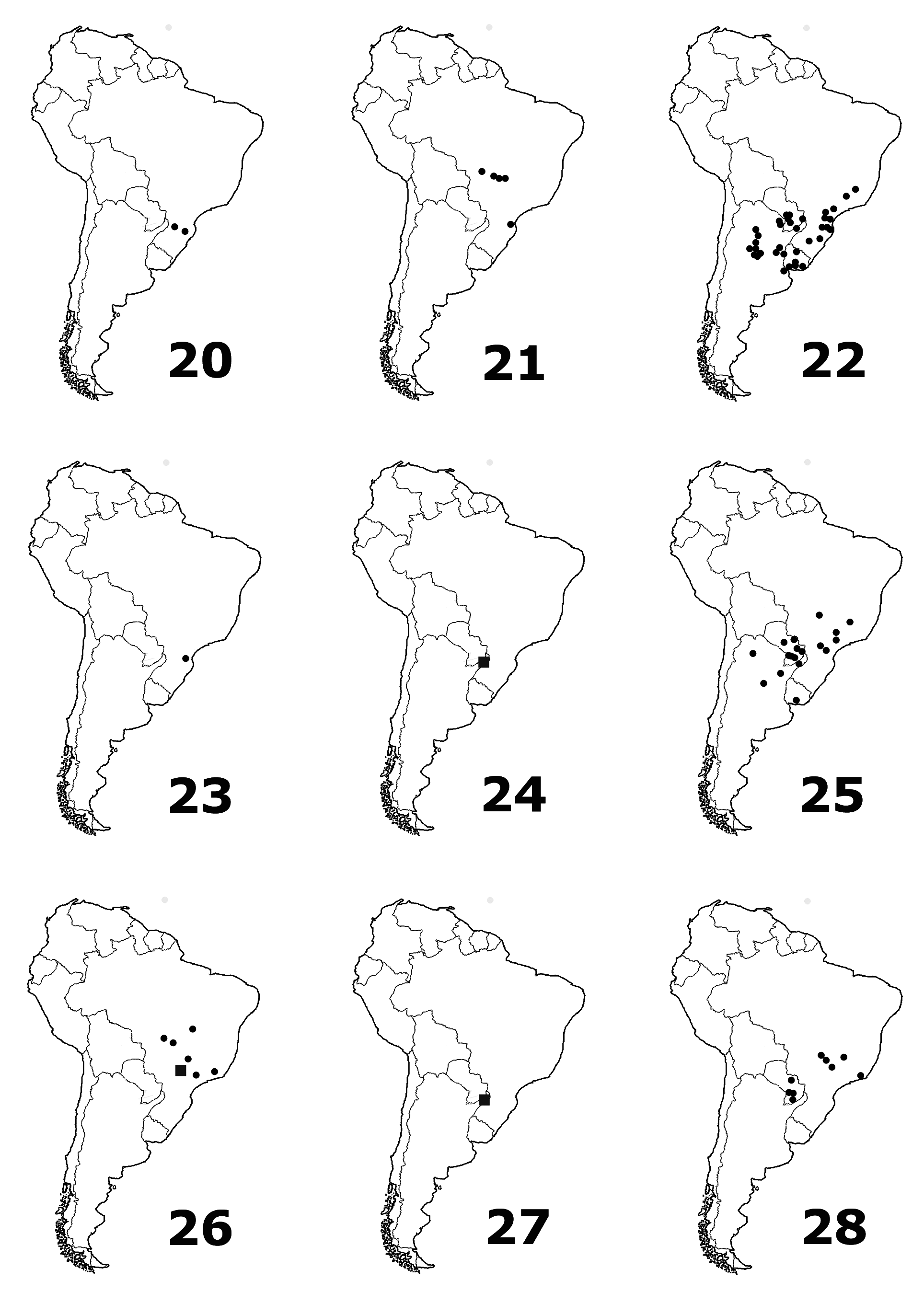
Further material examined. BRAZIL: Lagoa Santa Reinhardt (1, ZMUC) ; São Paulo M. Ráz (3, NHMP) ; São Paulo C. Jordaõ 12.X.1959 G. Halik (1, USNMNH) ; Paranà Vila Velha 12.I.1969 C. W. O'Brien (2, BYU) ; Jataí Goiás (1, USNMNH) ; Paranà Ponta Grassa II.1947 Justus (1, USNMNH) ; Rio Grande (1, BMNH) ; Minas Geraes Pocinhos 7.X.1935 (1, USNMNH) ; Campos do Tobagy v. Königswald S Angek 2.X.1904 (1, MNHUB) ; “ Brésil ” (1, USNMNH) ; “ Brasilia ” (1, ZMUC) ; “ Brasilia ” coll. H. Clavareau 1932 (1, MNHN) ; “ Brésil ” (6, MHNH) . PARAGUAY: Colon Risso Dr. Ternez (1, NHMB) ; Sapucay W. T. Foster (1, USNMNH) ; Dept. Amambay Parq. Nac, Cerro Cora 24.II.1981 (1, USNMNH) ; San Bernardino W. Eisenlehr (1, MNHUB) ; San Bernardino K. Fiebrig (4, USNMNH) ; San Bernardino XI.1898 G. Boggiani (2, MSNG) ; Caaguazú XI.1950 Reitter (4, NHMB) ; Canguazu [Caaguazú] XI.1950 (1, NHMB) ; Kanindeyu Curuguaty 17.X.1991 U. Drechsel (2, MSPC) ; Itapua Vega XII.1954 (2, MNHUB) ; Paraguay J. & Tr. Šedý (1, NHMP) ; Paraguay C. Höge 26.II.1908; (1, NHMB) ; Paraguay C. Fiebrig (8, MNHUB) ; Asuncion Revoil 1891 (1, MNHN) . ARGENTINA: Chaco de Santa-Fé Las Garzas Bords du Rio Las Garzas 25 kil. a l'O d'Ocampo E.R. Wagner 1903 (1, MNHN) ; Territ. de Misiones Rio Parana (1, MNHUB) ; Misiones Pindapoy Monrós (1, USNMNH) ; “ Emb. Uruguay ” Misiones (1, MHNH) ; Cordoba Balneatia (1, MSNM) ; Salta (1, MNHUB) ; Salta Crestón de Metán II.1945 Monrós (1, USNMNH) . URUGUAY: Canelones, 19 Km NE Canelones R 63 7.II.1989 C. & L. O'Brien & G. Wibmer (1, BYU) .
Distribution. Argentina, Brazil, Paraguay, Uruguay. New to Argentina, Paraguay, and Uruguay.
Diagnosis. A Metallactus of medium size, particularly similar in shape and coloration to M. albipes . As a matter of fact, the size is the only “traditional” character that can help in allocating specimens to its proper species. However, the shape of sclerite IV is decisive in distinguishing male specimens from M. albipes , being the apex of ascending branches regularly rounded in M. corruptus and saddled in M. albipes . Coloration is strongly variable in both species, and also in M. corruptus it is possible to come across specimens with even a “ nigrofasciatus ” look. However, in M. corruptus lateral thickenings on sides of aedeagal apex are constantly lacking, but the more effective differential trait is the shape of the sclerite IV of the endophallus. M. guarani looks also very similar, but black pattern on elytra is generally less developed and, most importantly, the coloration of the abdominal ventrites is completely yellow (always partly black in M. corruptus ), besides, the aedeagus differs in having two elongate swellings, feeble but perceptible, on sides of aedeagal basal foramen.
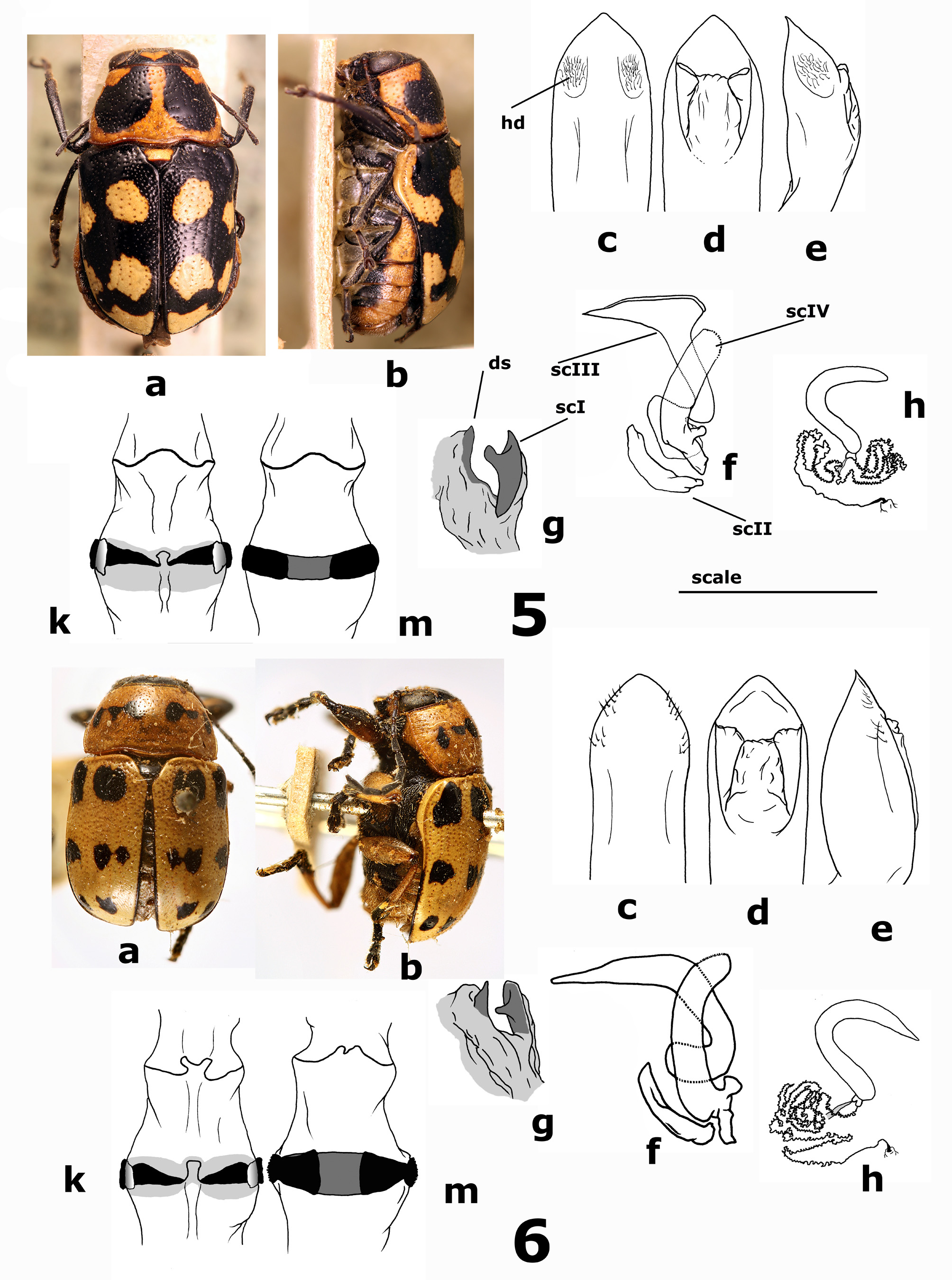
Description of male. Habitus in Fig. 6 View FIGURES 5–6 a–b (LT). BL = 4.7–5.7 mm, BW = 2.9–3.4 mm, PL = 1.4–1.9 mm, PW = 2.6–3.1 mm. Interocular distance 10.6–12.9% of BL.
Head with frons, clypeus and labrum yellow. Vertex, insertion of antennae and ocular canthus black. Black pattern can be more or less extended. Surface faintly setose with moderately impressed scattered punctures, but in some specimens punctures completely obliterated. Antennomeres 3 to 5 cylindrical, slender, barely setose, rather lustrous. The subsequent dull, weakly flattened.
Pronotum yellow with a median transverse wavy black stripe not reaching lateral margins. Such stripe can be reduced, split, turned into two/four little rounded spots or completely obliterated. Pronotal outline short and transverse, with narrow regularly curved lateral margins. Posterolateral impressions scarcely impressed so that posterior margin not or barely swollen. Surface moderately shiny with scattered medium impressed punctation.
Scutellum black, moderately raised with apex truncated or slightly rounded. Surface minutely punctured.
Elytra with “standard color pattern” as follows: surface yellow with two transverse black stripes, the first close to basal margin, the second just in the middle, two additional little black spots on apical clivus. Stripes often split in two rounded spots, giving a black pattern of 2+2+2 ( Fig. 6b View FIGURES 5–6 ). Black pattern can also be more or less obliterated. In some cases, elytra are completely yellow. Parallel-sided, regularly convex with base and middle equally wide. Surface even with inner side of humeral calli weakly impressed and post-scutellar area faintly raised. Surface moderately shiny with small and shallow punctures arranged in irregular rows, disappearing on elytral apex. Intervals smooth.
Pygidium yellow, smooth, haired, shallowly and sparsely punctured.
Inferior parts of thorax blackish with mesothorax often mostly yellow. Abdominal ventrites prevalently yellow on sides, often darkened towards median line. Prosternal process moderately broad, longitudinally sulcated with surface coarsely punctate, bearing long whitish setae. Legs mainly yellow, more or less infuscate in particular on tarsi, distal half of tibiae and fore femora.
Fifth abdominal ventrite with shallow median depression, triangular and short, barely reaching the mid of sternite. Median lobe of aedeagus ( Fig. 6 View FIGURES 5–6 c–e) very simply shaped, cylindrical, with smooth surface without sculpture or pitting and apex scarcely differentiated. Hairy dents almost obliterated.
Endophallus ( Fig. 6 View FIGURES 5–6 f–g) with sclerite I large, well sclerotized and pigmented, denticle prominent and dorsally projecting, dorsal spicule rather large, conical, pointed and pigmented. Sclerite II well developed, regularly bent at base. Apex of sclerite III fairly tapered and pointed. Branches of sclerite IV subequal or slightly shorter than sclerite III in the folded up structure, long, arched, weakly tapered towards apex, faintly bent at base, with crenulate upper rim and surface covered by tiny oblique wrinkles. In some Argentinian specimens sclerite IV is stouter, sclerite II is shorter and more abruptly bent at base, denticle of sclerite I is shorter, stouter and placed perceptibly further down on sclerite surface. Taxonomic significance of these differences, if any, is unclear.
Female. BL = 5.9–6.2 mm, BW = 3.5–3.8 mm, PL = 1.9–2.1 mm, PW = 3.0– 3.5 mm. Interocular distance 11.9–16.1% of BL.
Fifth abdominal ventrite in females with a rounded and quite deep pit. Vasculum of spermatheca ( Fig. 6h View FIGURES 5–6 ) scarcely pigmented, compact, with short upper branch, and apex gradually tapered, almost straight. Basal branch of vasculum almost straight. Ampulla fairly pigmented, slightly shifted on dorsal side of vasculum. Duct of uniform size, slender, coiled. Distal not coiled portion quite short. Insertion on bursa copulatrix lengthened and slightly swollen, clearly pigmented. Rectal apparatus ( Fig. 6 View FIGURES 5–6 k–m) with dorsal sclerites laterally not projected beyond rectus, narrow, tapered towards median line, with apodemes quite wide, hyaline but clearly perceivable, bent upwards and leaning against rectum. Ventral sclerite large, ribbon-like, generally tapered on sides, more or less pigmented in middle, with rounded rather short apodemes laterally projecting.
Remarks. The species is definitely very variable in terms of dorsal coloration. Starting from the typical color habitus, dorsal black pattern can be almost completely obliterated or significatively more developed. It cannot be excluded that some of the chromatic forms might characterize genetically distinct populations. For instance, some Paraguayan light colored specimens seem to have antennae slightly stouter than the Brazilian ones, and thus may have some interest from a taxonomic and biogeographic point of view. However, the great variability and the lacking of significative series of specimens do not allow so far to define in a non-ambiguous way any taxonomically coherent groups within this species.
| MLUH |
Martin Luther Universitaet |
| NMV |
Museum Victoria |
| WB |
Department of Bacteriology, University of Wisconsin |
| ZMUC |
Zoological Museum, University of Copenhagen |
| BYU |
Monte L. Bean Life Science Museum |
| MNHN |
Museum National d'Histoire Naturelle |
| NHMB |
Natural History Museum Bucharest |
| MSNG |
Museo Civico di Storia Naturale di Genova 'Giacomo Doria' |
| MSPC |
Museo di Storia Naturale "Pietro Calderini" |
| MSNM |
Museo Civico di Storia Naturale di Milano |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Cryptocephalinae |
|
Genus |
Metallactus corruptus Suffrian, 1866
| Sassi, Davide 2018 |
Metallactus corruptus
| Blackwelder, R. E. 1946: 641 |
| Clavareau, C. H. 1913: 92 |
| Suffrian, E. 1866: 370 |