
Lacunicambarus dalyae Glon, Williams and Loughman, 2019
|
publication ID |
https://doi.org/10.11646/zootaxa.4683.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:13A59FF8-C1A1-4026-BD1F-ED83F859B539 |
|
persistent identifier |
https://treatment.plazi.org/id/11D69719-EFEA-46B8-A9D2-506FF0F5B494 |
|
taxon LSID |
lsid:zoobank.org:act:11D69719-EFEA-46B8-A9D2-506FF0F5B494 |
|
treatment provided by |
Plazi |
|
scientific name |
Lacunicambarus dalyae Glon, Williams and Loughman |
| status |
sp. nov. |
Lacunicambarus dalyae Glon, Williams and Loughman View in CoL sp. nov ( Figures 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 , 6 View FIGURE 6 )
Cambarus ( Tubericambarus) sp. A—Jezerinac 1993: 535 [in part].
Cambarus sp. A—Taylor & Schuster 2004: 126 [in part].
Cambarus polychromatus —Taylor et al. 2007:383 View in CoL [in part]. Schuster et al. 2008:502. Simon 2011:77 [in part]. Smith et al. 2011:38. Miller & Stewart 2013:270.
Cambarus ( Tubericambarus) polychromatus —Taylor & Schuster 2007:8 View in CoL . Miller & Stewart 2013:276. Miller et al. 2014:15.
Cambarus ( Tubericambarus) View in CoL sp. B—Heath et al. 2010:150.
Cambarus diogenes — Helms et al. 2013:1333 View in CoL [in part]. Clay et al. 2017:1177.
Cambarus ( Lacunicambarus) erythrodactylus — Simon et al. 2015:576 View in CoL [in part]
Lacunicambarus aff. polychromatus — Glon et al. 2018:604 View in CoL [in part]. Glon et al. 2019:456 [in part].
Diagnosis.— Eyes pigmented, not reduced. Rostrum narrow, moderately deflected, curving downward in lateral view, margins moderately thickened to acumen, lacking marginal spines or tubercles and median carina, shallowly excavated. Acumen distinctly delimited basally by 45° angles. Cephalothorax subcylindrical, laterally compressed, with 1–10 (mean: 6) adpressed tubercles lining posterior margin of cervical groove. Suborbital angle acute. Postorbital ridges developed, ending cephalically in small tubercle. Areola obliterated, constituting in adults 40–45% (mean: 42%) of entire length of cephalothorax. Antennal scale 2.7 to 3.6 (mean: 3.0) times as long as wide, broadest at mid-length, antennal spine strongly developed. Dorsomesial 1/4–1/3 surface of palm of chela studded with distinct to adpressed tubercles, mesial-most row consisting of 6–9 (mean: 8). Opposable margin of dactyl with concavity just proximal to midpoint. Ratio of dactyl length to palm length 1.6–2.0 (mean: 1.8). Dorsomesial longitudinal ridges of dactyl and propodus weakly developed. Dorsolateral impression at base of propodus weak. Ventral surface of chela with 0–7 (mean: 1) subpalmar tubercles. Mesial ramus of uropod with distomesial spine not reaching caudal margin. Gonopods of Form I males contiguous at base, with pronounced umbo near mid-length of caudal surface; terminal elements consisting of 1) long central projection lacking subapical notch, slightly tapered at mid-length, distally rounded, slightly shorter than mesial process, directed caudally at approximately 90 o, overreaching margin of umbo by noticeable amount, 2) long mesial process with conical base, tapering slightly near mid-length, tipped with protruding finger, directed caudally at approximately 90 o and overreaching margin of umbo by noticeable amount, 3) inconspicuous caudal knob protruding from caudolateral base of central projection. Hook on ischium of third pereiopod only. Female with annulus ventralis subcircular or subquadrangular, slightly wider than long, deeply embedded in sternum, with anterior half mildly pliable and posterior half sclerotized.
Holotypic male, Form I (Catalogue # OSUMC 10855; Figures 1 View FIGURE 1 , 2 View FIGURE 2 A–D, G–I, K, 6; Table 2 View TABLE 2 ).— Cephalothorax bullet-shaped in dorsal view ( Figure 2G View FIGURE 2 ), width 93% of depth. Abdomen narrower than cephalothorax (19.1 and 22.5 mm, respectively; Figure 1 View FIGURE 1 ); maximum width of cephalothorax less than depth at caudodorsal margin of cervical groove (22.5 and 24.3 mm, respectively). Areola obliterated; length 42% of total length of cephalothorax ( Figure 2G View FIGURE 2 ). Rostrum moderately deflected and curved downwards in lateral view, margins slightly thickened; acumen distinctly delimited basally by 45° angles, anterior tip upturned, not reaching ultimate podomere of antennular peduncle; dorsal surface of rostrum shallowly concave with minute punctations forming single row along margins. Subrostral ridge moderate, evident in lateral aspect along entire length of rostrum. Postorbital ridges developed, grooved dorsolaterally, ending cephalically in small tubercle. Suborbital angle acute; branchiostegal spine absent ( Figure 2A View FIGURE 2 ). Posterior margin of cervical groove lined by collar of 9 adpressed tubercles. Branchiostegal region granulate. Anteroventral branchiostegal region with 14 small tubercles. Hepatic region with scattered granules and tubercles. Remainder of cephalothorax with slight punctations dorsally and laterally. Abdomen longer than cephalothorax (52.4 and 50.5 mm, respectively), 2.7 times as long as wide; pleura short, rounded caudoventrally or ventrally. Cephalic section of telson with 2 spines in caudolateral corners, mesial spine slightly moveable. Proximal segment of lateral ramus of uropod with border of 17 spines on distal margin, second-most lateral spine distinctly longer than others; mesial ramus of uropod with prominent median rib ending distally as strong distomesial spine not reaching margin of ramus, laterodistal spine of ramus strong.
Cephalomesial lobe of epistome ( Figure 2I View FIGURE 2 ) bell-shaped with uniform raised margins, ventral surface shallowly concave; main body of epistome with shallow fovea; epistomal zygoma arched. Ventral surface of antennular peduncle’s proximal podomere with small spine at mid-length. Antennal peduncle without spines. Antennal scale 3.0 times as long as wide ( Figure 2H View FIGURE 2 ), broadest near mid-length, lateral margin straight from basal area to broadest distal point, ending in strongly developed antennal spine reaching past proximal margin of ultimate podomore of antennal peduncle. Ventral surface of entire third maxilliped densely studded with long, flexible setae; distolateral angle not acute.
Length of right chela ( Figure 2K View FIGURE 2 ) 89% of cephalothorax length; chela width 46% of chela length; palm length 33% of chela length; dactyl length 1.9 times palm length. Dorsomesial 1/3 surface of palm of chela studded with distinct to adpressed tubercles, most strongly developed mesially, mesial-most row consisting of 7; proximal dorsolateral half smooth, distolateral area punctate, punctations deep, large in vicinity of dorsolateral base of propodus; lateral margin of propodus not costate; ventromesial surface of palm with small punctations, 2 bulbous tubercles on articular rim opposite base of dactyl, lateral-most spiniform; 1 subpalmar tubercle. Both fingers of chela with weakly developed dorsomedian longitudinal ridges. Opposable margin of propodus with row of 9 tubercles, decreasing in size from base except for 3 rd from base which is greatly enlarged over adjacent tubercles, ultimate tubercle with corneous tip, larger than penultimate tubercle, positioned ventrally relative to adjacent tubercles; single row of minute denticles extending distally from fifth tubercle. Opposable margin of dactyl with row of 12 tubercles decreasing in size from base except for 3 rd from base which is greatly enlarged over adjacent tubercles; conspicuous gap between 2 nd and 3 rd tubercles; single row of minute denticles extends distally from 7 th tubercle; mesial surface of dactyl studded with 15 tubercles proximally, not forming distinct rows, giving way to punctations distally. Dorsolateral impression at base of propodus weak.
Cheliped carpus with distinct dorsal furrow; dorsomesial surface with 5 scattered tubercles; dorsolateral surface punctate; mesial surface with 5 scattered tubercles plus 1 large procurved spine near distal margin; ventral surface with spine on distal articular rim. Merus with 2 pre-marginal spines dorsally, ventrolateral margin with row of 3 spines, ventromesial margin with row of 11 spines, increasing in size from base. Basioischial segment of first pereiopod with 3 small tubercles on ventral margin. Ischium of 3 rd pereiopod with simple hook extending proximally to basioischial margin, not opposed by tubercles on basis. Coxa of 4 th pereiopod with setiferous, caudomesial boss, ventral surface calcified; coxa of 5 th pereiopod lacking boss, ventral surface membranous.
Gonopods contiguous at base, reaching past caudomesial boss of 4 th pereiopod; terminal elements as described in diagnosis ( Figure 2 View FIGURE 2 B–D).
Dextral gonopod and antennal scale separated from specimen and placed in vials inside specimen jar. One gill has been extracted from sinistral gill chamber of specimen, preserved in 100% ethanol and frozen at OSUMC for future DNA extractions (MGG 668).
Allotypic female (Catalogue # OSUMC 10857; Figure 2J View FIGURE 2 ; Table 2 View TABLE 2 ).— The allotypic female differs from the holotype as follows: Areola length 43% of total length of cephalothorax. Anterior tip of rostrum acumen overreaching proximal margin of ultimate podomere of antennular peduncle. Posterior margin of cervical groove lined by collar of 6 adpressed tubercles. Anteroventral branchiostegal region with 11 small tubercles. Proximal segment of lateral ramus of uropod with border of 16 spines on distal margin. Antennal scale 2.9 times as long as wide. Length of right chela 72% cephalothorax length; chela width 48% of length; dactyl length 1.4 times palm length. Mesial-most row of tubercles on dorsomesial surface of palm of chela consisting of 8 tubercles, chela with 2 subpalmar tubercles. Opposable margin of dactyl with row of 15 tubercles, decreasing in size from the 5 th tubercle; no conspicuous gap between tubercles of proximal half. Mesial surface of dactyl studded with 17 tubercles, decreasing in size distally. Dorsomesial surface of cheliped carpus with 7 scattered tubercles; mesial surface with 5 spiniform tubercles plus 1 large procurved spine near distal margin. Merus with 5 pre-marginal spines dorsally, 2 of which are considerably larger than others; ventrolateral margin with row of 4 spines.
Annulus ventralis ( Figure 2J View FIGURE 2 ) as in diagnosis; 1.3 times wider than long, with bifurcated leathery ridge mesially located in anterior half; tongue extending from sclerotized lingual (sinistral) wall into fossa of sclerotized supralingual (dextral) wall; supralingual wall more swollen caudomesially than lingual wall, both walls curved on outer margin. Posterior margin of annulus ventralis reaching anterior margin of oblong, approximately symmetrical postannular sclerite, lacking setae. First pleopods overreaching distal edge of annulus ventralis when abdomen flexed.
Morphotypic male, Form II (Catalogue # OSUMC 10856; Figure 2 View FIGURE 2 E–F; Table 2 View TABLE 2 ).— The morphotypic Form II male differs from the holotype as follows: cephalothorax width 99% of depth; areola length 43% of total length of cephalothorax. Anterior tip of rostrum acumen overreaching proximal margin of ultimate podomere of antennular peduncle. Posterior margin of cervical groove lined by collar of 6 adpressed tubercles. Anteroventral branchiostegal region with 8 small tubercles. Abdomen 2.6 times as long as wide. Proximal segment of lateral ramus of uropod with border of 15 spines on distal margin. Antennal scale 2.6 times as long as wide; lateral margin slightly concave. Length of right chela 80% of cephalothorax length; palm length 34% of chela length; dactyl length 1.8 times palm length. Opposable margin of propodus with row of 8 tubercles. Opposable margin of dactyl with row of 9 tubercles. Mesial surface of dactyl studded with 14 tubercles. Dorsomesial surface of cheliped carpus with 7 scattered tubercles; mesial surface with 4 scattered tubercles plus 1 large procurved spine near distal margin. Merus ventrolateral margin with row of 2 spines, ventromesial margin with row of 10 spines.
Form II gonopod central projection non-corneous, rounded, slightly shorter than mesial process, overreaching margin of umbo ( Figure 2 View FIGURE 2 E–F). Mesial process conical at base, tapering to a thin tip, overreaching margin of umbo. Caudal knob not visible. Dextral gonopod separated from specimen, placed in glass vial inside specimen jar. Sinistral chela regenerated, extreme tip of dactyl of dextral chela damaged.
Coloration and color pattern. Like many cambarid crayfishes, L. dalyae sp. nov. exhibits a notable ontogenetic shift in its coloration and color pattern (MGG, personal observation).
Large adult specimens resemble the holotype and exhibit the most striking coloration found in this species ( Figure 1 View FIGURE 1 ). The background coloration of the cephalothorax and chela of these large specimens is predominantly golden. The dorsal surface of the cephalothorax is darker than the lateral surfaces, which are subtly mottled. The legs of large adults are a light amber or cream color. The background coloration of the abdomen is typically darker than the cephalothorax, ranging from ocher to bronze. The background color of the crayfish is complemented by polychromatic highlights throughout the body. The dorsal surface of the carpus and merus are highlighted in shades of turquoise and emerald. The surfaces of the telson and rami of clean (i.e., recently molted) specimens are blue with white speckles. Much of the crayfish is highlighted in a ruby red color, including the tips of the chela, the tubercles and spines of the chela, merus and carpus, and the margins of the antennal scales, rostrum, postorbital ridge, pereiopods 2–5, abdomen, telson and uropods. The dorsal side of the cephalothorax and abdomen is adorned by a single longitudinal gladiate stripe that is typically yellow. Lastly, the dorsal side of the cephalon is adorned by a diamond-shaped pattern formed anteriorly by the ruby red highlights of the rostral margins and postorbital ridge and posteriorly by two ruby red to orange diagonal lines running from the distal ends of the postorbital ridge to a convergence point just anterior to the cervical groove.
The coloration and color pattern of young adult specimens differs slightly from adults ( Figures 3 View FIGURE 3 , 4 View FIGURE 4 ). Firstly, these specimens exhibit more pronounced mottling on their cephalothoraxes and abdomens. In addition, the background coloration of these younger specimens is less vibrant, ranging from burnt orange to olive. These specimens have more conspicuous turquoise and emerald highlights on their merus, carpus and chela than large adults. Lastly, the diamond-shaped pattern on the cephalon is sometimes less clearly delineated in these specimens, but it is typically still present.
Juvenile specimens are the darkest and most mottled specimens of the species. The background color of their entire body including the chela ranges from bronze to brown. They have conspicuous mottling on the lateral surfaces of the abdomen and cephalothorax which is of a muted goldenrod color. These specimens still bear characteristic markings of the species, including the dorsal longitudinal gladiate stripe, which is also a goldenrod color, and the diamond-shaped pattern on the cephalon. Lastly, these specimens have similar, albeit more muted, ruby red highlights on the margins of somites throughout the body.
Size. The largest L. dalyae sp. nov. that we have examined is a Form I male from Prentiss County, Mississippi that measures 53.4 mm carapace length (CL). The average ± standard deviation size of specimens that we examined was 43.0 ± 5.2 mm CL for Form I males, 38.4 ± 4.5 mm CL for Form II males, and 42.4 ± 3.5 mm CL for Females. The smallest Form I male that we examined measured 33.9 mm CL; however, Miller et al. (2014) recorded collecting a Form I male specimen measuring 27.4 mm CL.
Variation. There is a moderate amount of morphological variation across the range of L. dalyae sp. nov. We have examined specimens from Tennessee with lanceolate rostrums lacking clearly delimited acumens and others with rostral margins that remain thickened and entire to the tip of the acumen. One specimen from Pike County, Alabama has distinctly concave rostral margins. Some specimens from across the range have a very slight depression at the tip of the central projection that one might label as a subapical notch, but we hesitate to do so as this feature is considerably more subtle than the deep and conspicuous subapical notch found in some other members of the genus (see key below). We have also seen variation across the species’ range in the distal end of the mesial process, which is occasionally tipped with one or more minute, finger-like tubercles, sometimes oriented laterally creating a fork. In some specimens, the lateral-most of the 2 bulbous tubercles on the articular rim opposite the ventral base of the dactyl is topped with a sharp spine oriented distally. One specimen from Lee County, Georgia exhibited an impressive and highly unusual 23 subpalmar tubercles, and a specimen from Walton County, Florida bore uncharacteristically long and thin chelae that did not appear to have been regenerated. Lastly, two specimens from Davidson County, Tennessee have remarkably thin antennal scales.
As mentioned above, much of the color variation found in L. dalyae sp. nov. is ontogenetic in nature; however, there appears to be a fair amount of color variation across this species’ range. Particularly noteworthy are specimens from Alabama whose background colors are bright green or orange, rather than the usual gold (Guenter A. Schuster, personal communication). Despite this color variation, these specimens were still recognizable by their longitudinal gladiate stripe and diamond-shaped pattern on the cephalon, suggesting that the color pattern of this species is more characteristic than the color itself.
Disposition of Types. The holotype, morphotype and allotype are deposited in the OSUMC (OSUMC 10855– 10857). Paratypes are deposited in the NCSM (NCSM 90130–90132).
Type locality. We collected the holotype and allotype from the banks of Big Richland Creek, a tributary of the Tennessee River in Humphreys County, Tennessee ( 36.1576° N, 87.8203° W). Most of the burrows at this site were within 1 meter of the stream channel and relatively shallow. These burrows were surrounded by dense ground vegetation and occasional young trees. The soil at this site was primarily clay mixed with a small amount of sand and pebbles.
We collected the morphotype and paratypes from burrows in and above the floodplain of Hurricane Creek, a tributary to the Duck River in Humphreys County, Tennessee ( 35.9823° N, 87.8148° W). The burrows that we excavated at this site were up to 10 m away from the stream channel and surrounded by ground vegetation (including a large amount of poison ivy) and mature trees. The terrain beyond the floodplain of Hurricane Creek at this site is steeply sloped causing some burrow entrances to be over 2 m above the water table and therefore particularly difficult to collect crayfishes from. The soil at this site was mostly clay loam with some sand.
Range. This species is widely distributed across the southeastern United States, having so far been found in Alabama, Florida, Georgia, Mississippi and Tennessee ( Figure 5 View FIGURE 5 ). The western limit of its range appears to be the Mississippi River, as we are not yet aware of any records of L. dalyae sp. nov. from Louisiana, Arkansas or Missouri. The eastern limit of this species’ range is less clearly defined; we have numerous samples of this species from the Florida panhandle and the Upper Coastal Plain in Georgia, but further sampling will be required to more thoroughly determine its range in these states. Similarly, the northern extent of the species’ range is unclear. It is found in West and Middle Tennessee, but we have not yet located it in East Tennessee or Kentucky. Kentucky is inhabited by a number of other Lacunicambarus species, including the closely-related L. polychromatus , so it is possible that competitive exclusion has prevented L. dalyae sp. nov. from expanding its range into this State. Similarly, East Tennessee is occupied by L. aff. thomai ( Jezerinac, 1993) and L. acanthura ( Hobbs, 1981) , and this region is also characterized by an increase in topographic relief and associated change in hydrology, to which L. dalyae sp. nov. may not be adapted. Overall, this species’ range does not correspond cleanly with any major drainage basin or ecoregion, but most of its range is within the confines of the South-Atlantic Gulf River Basin and the Southeastern Plains and Interior Plateau regions of the Southeastern USA Plains (sensu Wiken et al., 2011).
Specimens examined. We examined a total of 175 specimens from 31 counties in five States. See Table 3 View TABLE 3 for information on these specimens.
Conservation status. Lacunicambarus dalyae sp. nov. is widely distributed throughout the southeastern United States and common in appropriate habitat. It appears to persist amidst a moderate amount of anthropogenic habitat degradation, as evidenced by specimens collected from burrows in roadside ditches and urban lawns. We suggest that it be considered Currently Stable following the American Fisheries Society’s Endangered Species Crayfish Subcommittee criteria (Taylor et al., 2007) and of Least Concern following the IUCN criteria ( IUCN, 2012), but also recommend that potential threats to this species, including from pet trade collectors, be preemptively examined.
Life history notes. A detailed life history of L. dalyae sp. nov. in Alabama has previously been published ( Miller et al., 2014). Amongst their findings, Miller et al., 2014 reported that mating of L. dalyae sp. nov. occurs in burrows during autumn or winter, with females extruding up to 171 eggs at the end of that same winter. Our examination of specimens largely corroborates this timing except for a single gravid female from Alcorn County, Mississippi that was collected on August 27 th, 1936. Overall, this phenology is similar to what has been documented for other Lacunicambarus species (e.g., Fitzpatrick, 1978; Thoma et al., 2005; Glon et al., 2019). In addition, Miller et al., 2014 found that juvenile L. dalyae sp. nov. are released and spend a period time in open water habitats before emerging to dig their first burrows in the banks or floodplain, which is a dispersal strategy that may contribute to the wide range of L. dalyae sp. nov. and other Lacunicambarus species ( Helms et al., 2013; Clay et al., 2017; Glon et al., 2018). Based on their length-frequency histograms, Miller et al., 2014 suggested that the minimum lifespan of this species is 4 years. We do not have a reliable way of aging our specimens, but we expect that the maximum lifespan of this species could be as much as twice this minimum lifespan estimate based on our observations of other Lacunicambarus species (e.g., Glon et al., 2019).
Ecological notes. Lacunicambarus dalyae sp. nov. is a primary burrowing crayfish typically found burrowing in alluvial soils of stream floodplains. Populations of this species are also frequently found burrowing in anthropogenically altered habitats like roadside ditches, the banks of ponds, and urban lawns. Like most of its congeners, this species excavates moderately complex burrows that consist of one or more surface openings ( Miller et al., 2014 found a range of 1–5 with a mean of 2.1) leading to tunnels that converge into a central tunnel approximately 30-50 cm below the surface. The depth attained by the central tunnel is largely dependent on the depth of the water table and substrate but can be greater than 2 m for adults. Depending on the time of year and weather conditions, burrow openings may be open or plugged, and one or more of these openings may be topped by a conspicuous chimney made of mud pellets excavated from the burrow.
Like its congeners and most other cambarid crayfishes, L. dalyae sp. nov. appears to be an opportunistic omnivore ( Reynolds et al., 2013). We have not collected dietary data for this species in situ but individuals that we have kept in captivity have thrived when fed a diet consisting of a mixture of foods including leaf detritus, live invertebrates, and shrimp- and spirulina-based commercial fish foods.

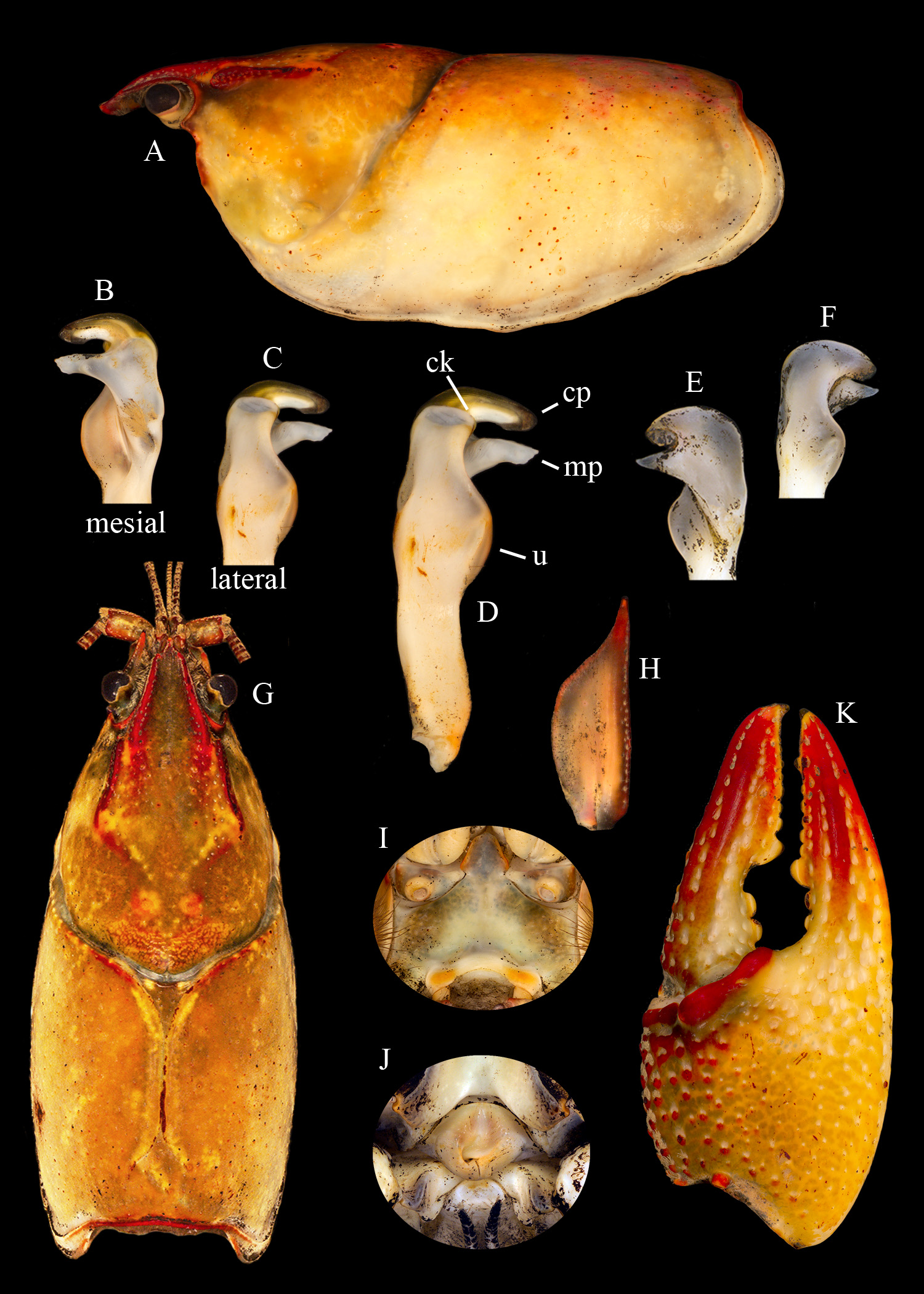


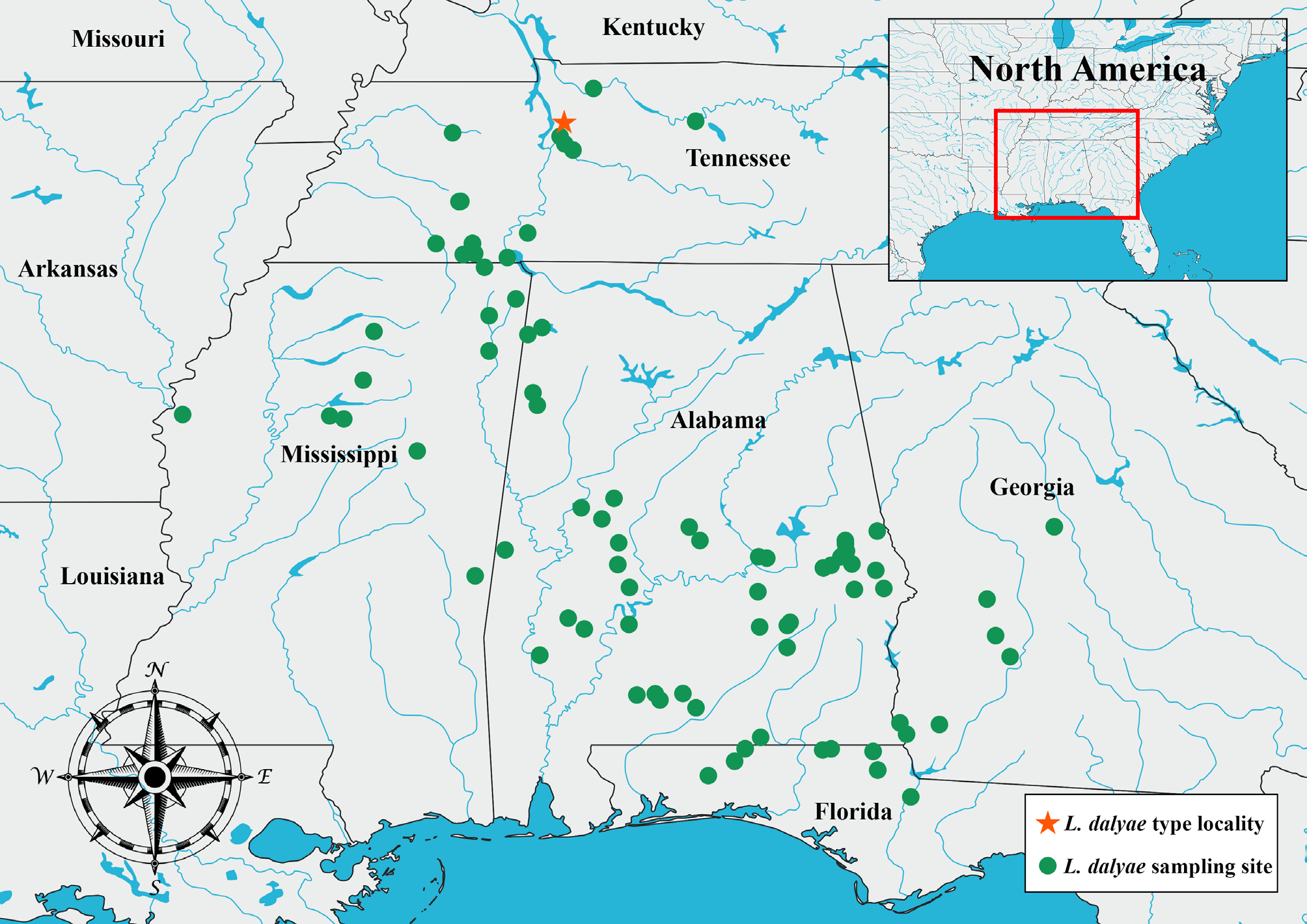
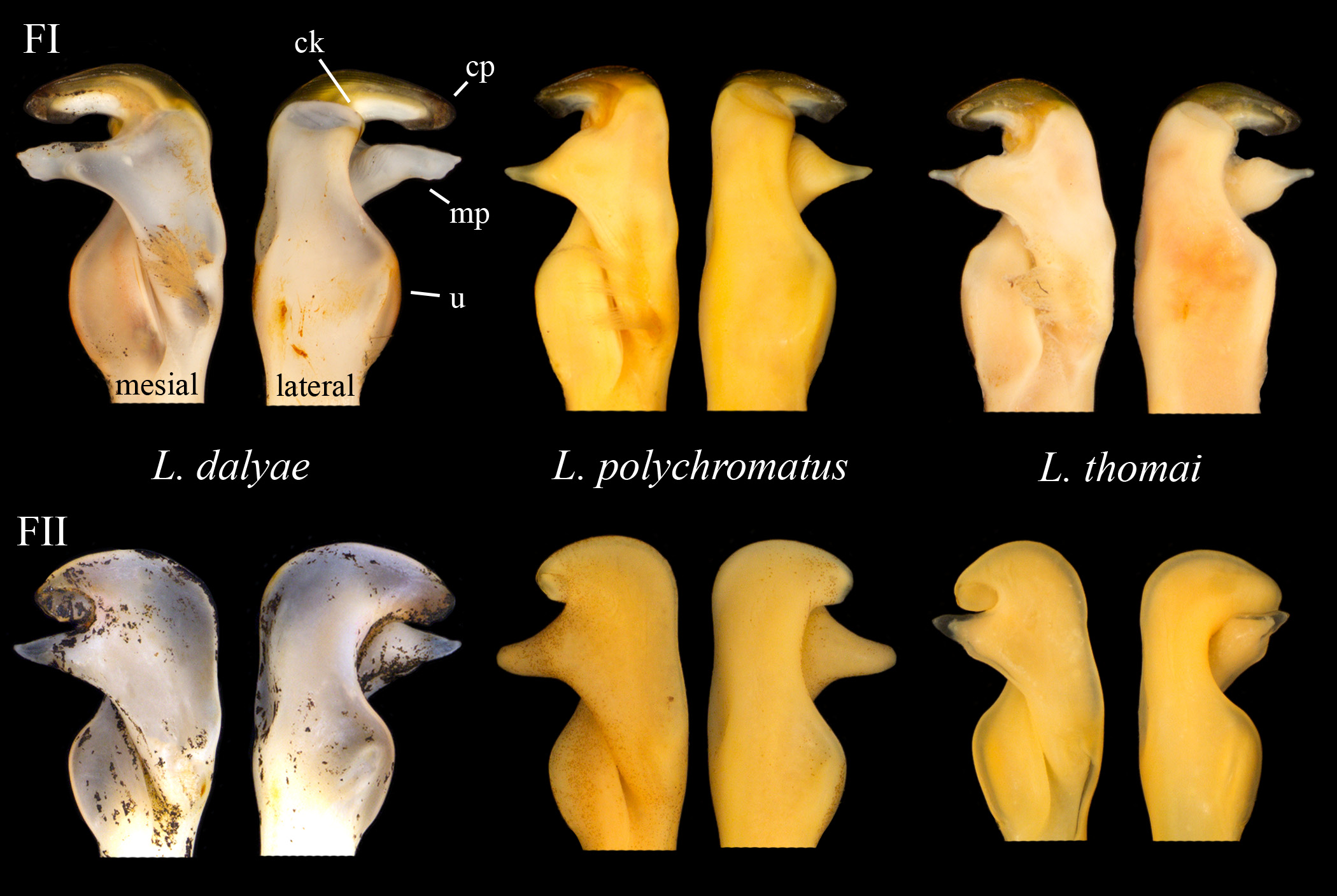
TABLE 2. Measurements (mm) of holotype, morphotype, and allotype of Lacunicambarus dalyae sp. nov. (OS- UMC 10855–10857).
| Character | Holotype | Morphotype | Allotype |
|---|---|---|---|
| Carapace: | |||
| Depth | 24.3 | 22.9 | 21.7 |
| Width | 22.5 | 22.6 | 20.2 |
| Length | 50.5 | 47.4 | 45.6 |
| Areola: | |||
| Length | 21.2 | 20.2 | 19.5 |
| Rostrum: | |||
| Width at eyes | 5.7 | 5.4 | 5.5 |
| Length | 8.2 | 7.6 | 7.6 |
| Postorbital ridge: | |||
| Width | 9.4 | 9.1 | 8.5 |
| Chela (right): | |||
| Length of propodus | 45.1 | 39.0 | 32.7 |
| Length of palm | 15.0 | 13.1 | 10.9 |
| Width of palm | 20.9 | 18.0 | 15.5 |
| Length of dactyl | 28.3 | 23.4 | 21.2 |
| Abdomen: | |||
| Length | 52.4 | 48.3 | 49.0 |
| Width | 19.1 | 18.4 | 18.1 |
| Gonopod: | |||
| Length | 12.0 | 11.8 | NA |
| Width at umbo | 3.2 | 3.0 | NA |
| Annulus ventralis: | |||
| Length | NA | NA | 3.3 |
| Width | NA | NA | 4.1 |
| Antennal scale: | |||
| Length | 7.3 | 6.4 | 6.7 |
| Width | 2.4 | 2.4 | 2.3 |
TABLE 3. Data (sorted by State and County) on examined specimens of Lacunicambarus dalyae sp. nov. including field/catalogue number, State and County of collection, GPS coordinates in decimal degrees, collection date, number of specimens of each sex and Form, whether sample included gravid females, and other crayfish species found at collection site.Asterisks denote type locality. Please note that some museum specimens were not associated with GPS coordinates so we estimated them based on provided locality information. Abbreviations: aff., affinis; AJ, Alan Jeon; AL, Alabama; AUM, Auburn University Museum of Natural History; C., Cambarus; DF, Daniel Folds; FL, Florida; GA, Georgia; L., Lacunicambarus; MGG, Mael Gianni Glon; MS, Mississippi; OSUMC; Ohio State University Museum of Biological Diversity Crustacean Collection; PM, Paul Moler; TN, Tennessee; USNM, Smithsonian National Museum of Natural History.
| Field/catalogue # | State | County | Latitude | Longitude | Collection date | Form I males | Form II males | Females | Gravid? | Juveniles Other species | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| USNM 130777 | AL | Chambers | 32.7734 | -85.2261 | 14/04/1970 | 1 | |||||
| Zac-181023-03 | AL | Clarke | 31.7447 | -88.0224 | 23/10/2018 | 1 | |||||
| USNM 146495 | AL | Conecuh | 31.4248 | -87.0655 | 11/04/1975 | 1 | yes | ||||
| USNM 209053 | AL | Conecuh | 31.3731 | -87.0266 | 27/07/1967 | 1 | 2 | ||||
| USNM 209315 | AL | Conecuh | 31.4145 | -87.2180 | 26/07/1967 | 1 | 1 | ||||
| USNM 145302 | AL | Conecuh | 31.4271 | -86.8364 | 6/04/1974 | 1 | 8 | ||||
| USNM 209047 | AL | Conecuh | 31.3085 | -86.7279 | 12/05/1968 | 1 | 2 | L. aff. diogenes | |||
| USNM 147256 | AL | Elmore | 32.5610 | -86.2085 | 12/08/1976 | 1 | 4 | ||||
| MGG-170830-01 | AL | Lee | 32.5010 | -85.4361 | 21/04/2017 | 2 | 2 | ||||
| AJ-4 | AL | Lee | 32.5929 | -85.4956 | 17/10/2018 | 1 | |||||
| USNM 144745 | AL | Lee | 32.6011 | -85.4877 | 12/05/1972 | 1 | 2 | 5 | 7 | ||
| USNM 144752 | AL | Lee | 32.6954 | -85.4903 | 8/03/1973 | 3 | |||||
| USNM 145290 | AL | Lee | 32.6674 | -85.4893 | 31/03/1973 | 2 | 4 | 5 | 4 | ||
| USNM 145198 | AL | Lee | 32.6082 | -85.4810 | 9/06/1973 | 2 | 1 | ||||
| USNM 145291 | AL | Lee | 32.5587 | -85.5273 | 12/03/1973 | 1 | 1 | L. aff. diogenes | |||
| AUM 22881 | AL | Macon | 32.4681 | -85.6719 | 26/03/2009 | 1 | |||||
| AUM 22903 | AL | Macon | 32.4681 | -85.6719 | 30/03/2011 | 1 | |||||
| AUM 23041 | AL | Macon | 32.4681 | -85.6719 | 25/08/2011 | 1 | |||||
| DF-3 | AL | Perry | 32.4959 | -87.3758 | 21/07/2018 | 1 | |||||
| USNM 130812 | AL | Pike | 31.8088 | -85.9724 | 16/04/1970 | 1 | |||||
| AJ-3 | AL | Russell | 32.2899 | -85.4154 | 10/10/2018 | 1 | |||||
| USNM 130789 | AL | Russell | 32.4478 | -85.2376 | 15/04/1970 | 1 | 1 | 1 | |||
| PM-8 | FL | Holmes | 30.9681 | -85.6079 | 18/03/2019 | 2 | yes | C. striatus | |||
| ...Continued next page | |||||||||||
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lacunicambarus dalyae Glon, Williams and Loughman
| Glon, Mael G., Williams, Bronwyn W. & Loughman, Zachary J. 2019 |
Lacunicambarus aff. polychromatus —
| Glon 2018: 604 |
Cambarus ( Lacunicambarus ) erythrodactylus —
| Simon 2015: 576 |
Cambarus polychromatus —
| Taylor 2007: 383 |
Cambarus ( Tubericambarus ) polychromatus —
| Taylor & Schuster 2007: 8 |
Cambarus diogenes — Helms et al. 2013:1333
| - Helms et al. 2013: 1333: 1333 |