
Homalium ovatifolium Appleq., 2018
|
publication ID |
https://doi.org/ 10.15553/c2018v731a4 |
|
DOI |
https://doi.org/10.5281/zenodo.6309028 |
|
persistent identifier |
https://treatment.plazi.org/id/03CAA927-3A13-FFBC-E757-F98AFC622E76 |
|
treatment provided by |
Carolina |
|
scientific name |
Homalium ovatifolium Appleq. |
| status |
sp. nov. |
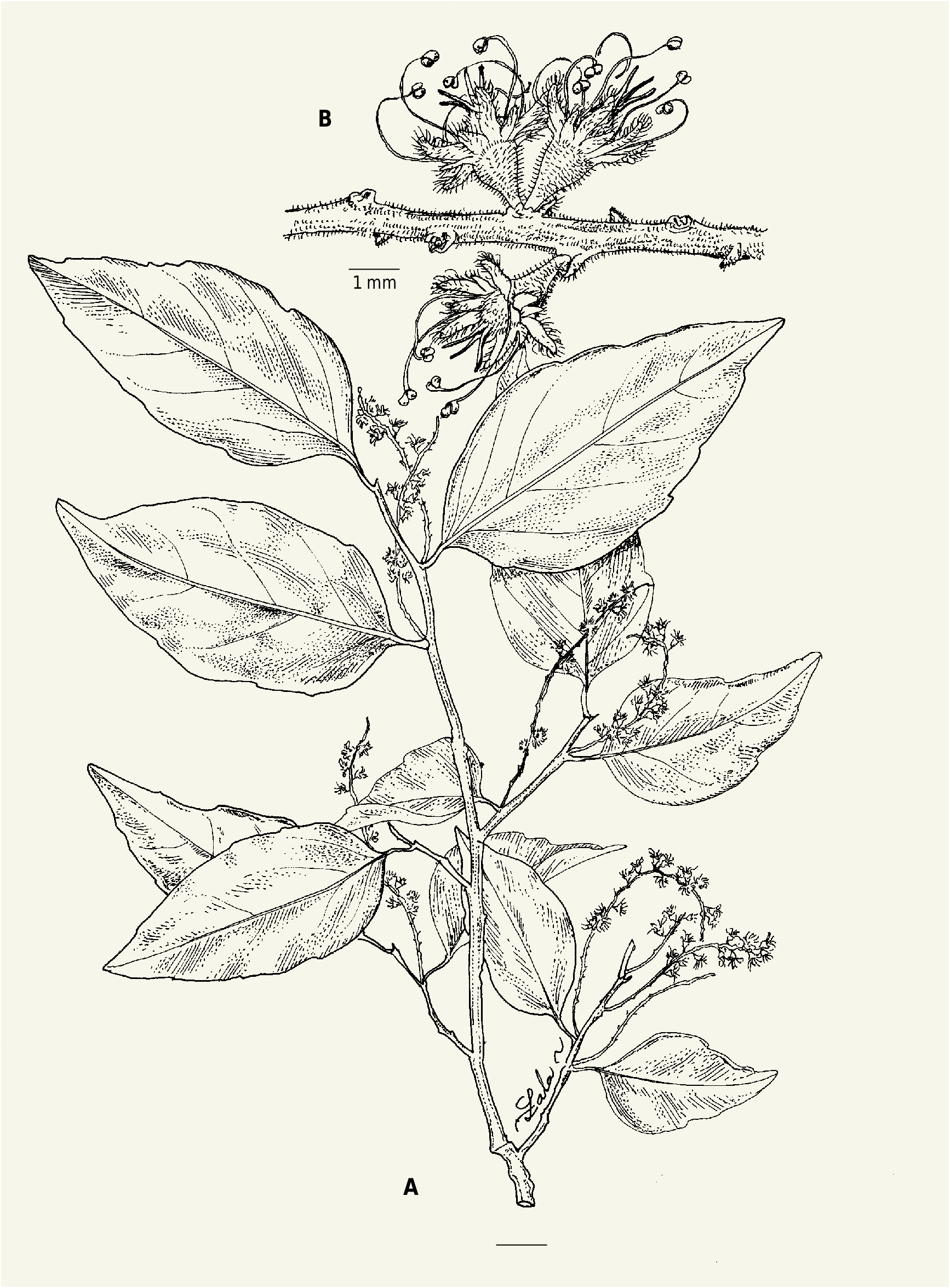
7. Homalium ovatifolium Appleq. View in CoL , spec. nova ( Fig. 4 View Fig ).
Holotypus: MADAGASCAR. Prov. Antsiranana: Ambilobe , Marivorahona , village le plus proche Betsimiranja , forêt d’Andohan’Antsohihy , Ambohibe, RN6 à 1 km au SE d’Ambilomagodra, 13°02’31’’S 49°09’19’’E, 100 m, 17.VII.2005, fl., Randrianaivo et al. 1206 (MO-6082912! GoogleMaps ; iso-: CNARP, P [ P04679126 ]!, TAN) .
Homalium ovatifolium Appleq. differs from H. longistaminum H. Perrier in having leaves larger, (4.2-)6.2-10.2 × (2.1-) 3.1-5.2 cm (vs (3.2-)3.5-7.5 × 1.6-3.3(-4.8) cm), ovate (to elliptical) (vs irregularly elliptical to broadly elliptical, ovate or rarely lanceolate) with the base rounded (vs convex to rounded), margins less toothed, sometimes subentire (vs irregularly crenate apically to crenate-serrate for most of their length), apex acute to acuminate (vs acute to obtuse or rounded, rarely shortacuminate), and petals narrowly oblong (vs oblong to obovate), the abaxial surface pubescent and the adaxial surface largely glabrous (vs both surfaces pubescent, the abaxial surface densely so and often the base of the adaxial surface densely so). It differs from H. masoalense Appleq. in having possibly broader leaves, ovate or rarely elliptical (vs often lanceolate), with the petiole longer, 5-11 mm long (vs 3-5.5 mm long); inflorescences shorter, (2.2-) 4-6.5 cm long (vs 5-11 cm long), often canescent (vs moderately pubescent), the flowers more often pedicellate with pedicels 0.5-1 mm long (vs absent or up to 0.5 mm long); sepals ovate to deltoid or narrowly deltoid, (0.3-)0.4-0.5(-0.7) mm long (vs oblong-lanceolate to narrowly oblong, 1.2-1.4 mm long); petals 0.8-1.2 mm long (vs 1.5-1.7 mm long), much longer than sepals (vs only moderately longer).
Candollea 73, 2018 Homalium sect. Odontolobus (Salicaceae) in Madagascar - 39
1 cm 1 cm 1 cm
Tree to 10 m tall, 50 cm dbh; young twigs glabrous. Leaves: petiole 5-11 mm long, glabrous; blade ovate (elliptical), (4.2-) 6.2-10.2 × (2.1-) 3.1-5.2 cm; base rounded; margins irregularly crenate-serrulate, sometimes with only 1 tooth per side, to slightly wavy or subentire; apex acute to acuminate. Inflorescences racemose, (2.2-) 4-6.5 cm long, canescent to moderately pubescent; flowers often in clusters of 2 or 3, pedicellate, with pedicels 0.5-1 mm long, or subsessile; bracts broadly ovate to deltoid or transversely oblong; bracteoles very thick, to subterete at base, densely pubescent, often caducous. Flowers 7-8-merous; sepals ovate to deltoid or narrowly deltoid, (0.3-) 0.4-0.5(-0.7) mm long; petals narrowly oblong, 0.8-1.2 mm long, margins long-ciliate, abaxial surface pubescent, mostly glabrous adaxially, apex rounded; filaments 1.4-1.9 mm long; upper surface of ovary densely pubescent to moderately so in fruit; styles 3-4, 1.3-1.8 mm long.
Vernacular name. – “Taindalitra” (Guittou et al. 160).
Distribution, ecology and conservation status. – Homalium ovatifolium is known from only two collections and a single location in deciduous forests at low elevation in extreme northern Madagascar in the western edge of the Andrafiamena Andavakoera protected area. Even if the only known location is within the protected area network, the new species is under threat and can be affected by a single event (e.g., fire), so its conservation status should be considered “Vulnerable” [VU D2].
Notes. – Homalium ovatifolium has clear affinities to H. longistaminum , which likewise has a canescent or mostly densely pubescent inflorescence, fleshy, densely pubescent bracteoles and long filaments and styles. It differs in its larger leaves, (4.2-) 6.2-10.2 × (2.1-) 3.1-5.2 cm (vs (3.2-)3.5-7.5 × 1.6- 3.3(-4.8) cm), which are more usually ovate with a rounded base, often long-acute to acuminate apex and few-toothed, shallowly wavy or subentire margins, and its narrow, less pubescent petals.
Interestingly, the two taxa are found at the opposite extremes of latitude, H. ovatifolium being confined to the extreme north of Madagascar and H. longistaminum to the southeast. It also is likely to be closely related to H. masoalense , which is from northeastern Madagascar and shares the unusual character of often ovate, acuminate leaves, as well as long filaments and styles. Homalium masoalense has often lanceolate, short-petiolate leaves and larger flowers (petals 1.5-1.7 mm long) with the sepals more similar to the petals in length (1.2- 1.4 mm long) and narrowly oblong to oblong-lanceolate.
Paratypus. – MADAGASCAR. Prov. Antsiranana: Ambilobe, Tanambao Marivorahona, Betsimiranja , Andohan’Antsohy , 4 km au NE de Betsimiranja, 13°02’32’’S 49°09’24’’E, 76 m, 2.VII.2005, fl. & fr., Guittou et al. 160 ( MO, P, TAN) GoogleMaps .
| CNARP |
CNARP |
| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
| TAN |
Parc de Tsimbazaza |
| MO |
Missouri Botanical Garden |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |