
Porrhomma convexum ( Westring, 1851 )
|
publication ID |
https://doi.org/10.11646/zootaxa.4481.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:BFC4982D-BB84-4141-BDFD-203F23CD1585 |
|
DOI |
https://doi.org/10.5281/zenodo.5963977 |
|
persistent identifier |
https://treatment.plazi.org/id/03C6A93B-FFC1-FFCF-FF7C-FB4885983C43 |
|
treatment provided by |
Plazi |
|
scientific name |
Porrhomma convexum ( Westring, 1851 ) |
| status |
|
Porrhomma convexum ( Westring, 1851) View in CoL
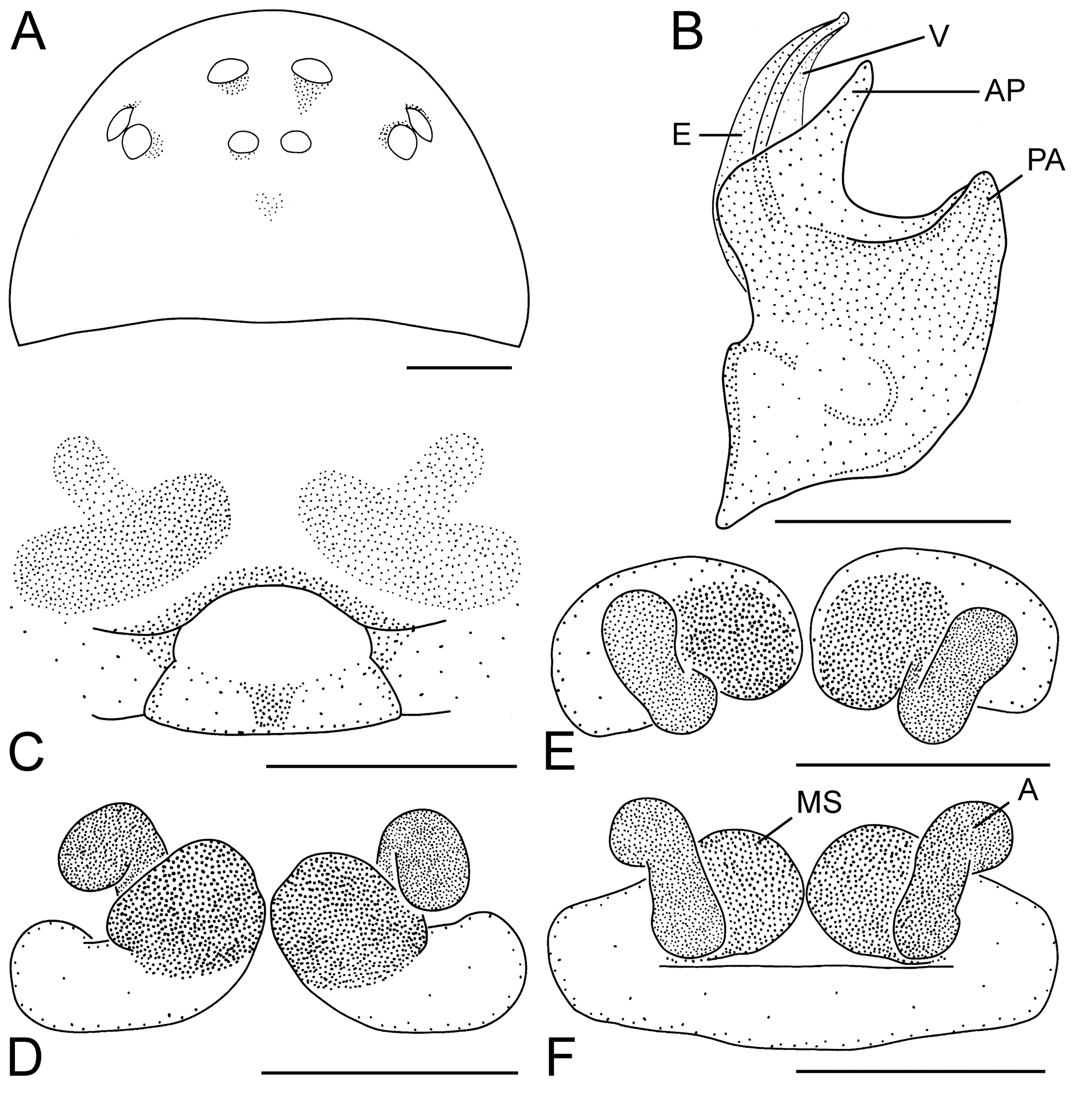
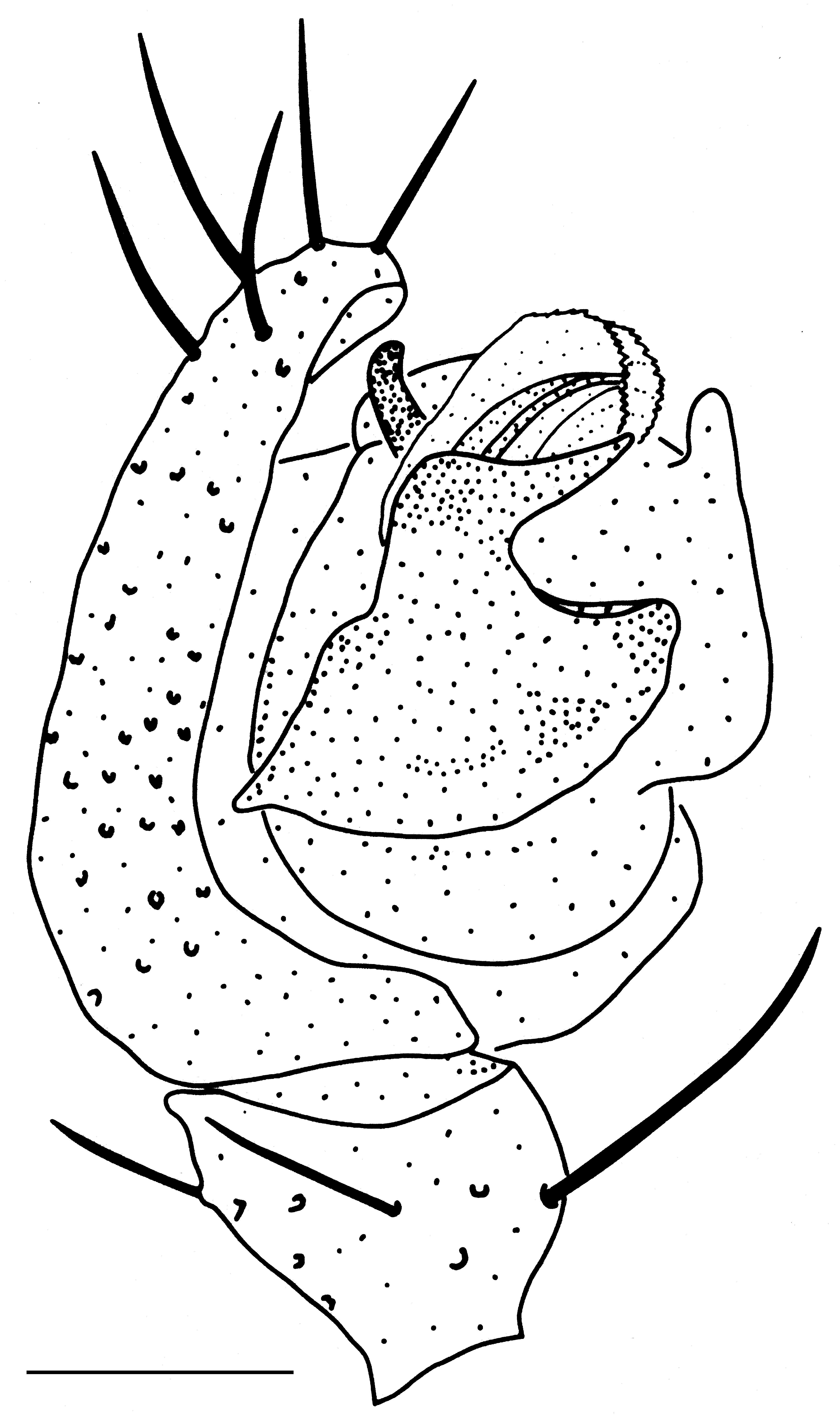
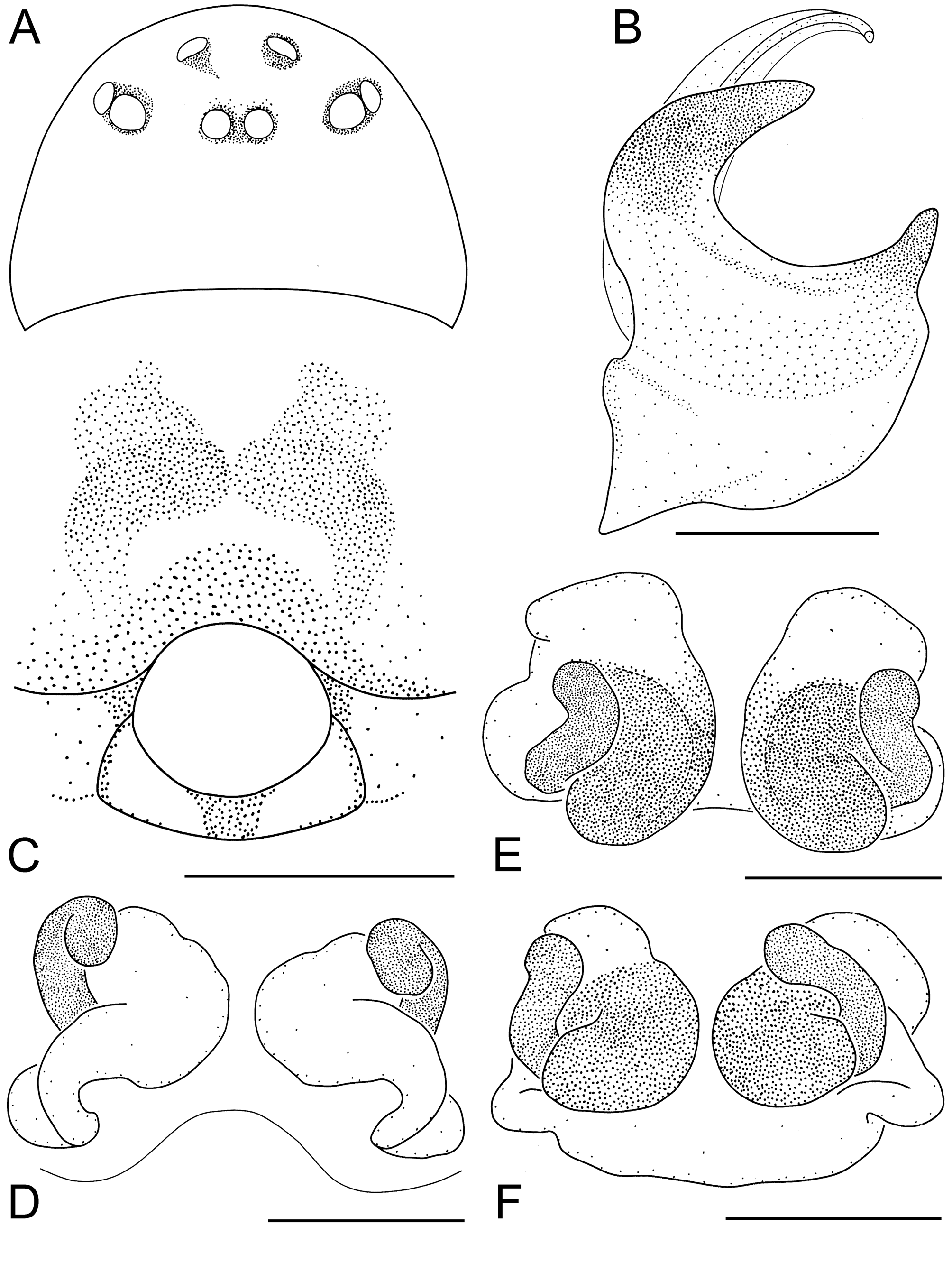
Figs. 19A–F View FIGURE 19 .
Erigone convexa Westring, 1851 — Westring (1851): p. 60 (descr. ♂ ♀); examined and transferred by Holm (1944).
E. reticulata Westring, 1851 — Westring (1851): p. 59; synonymised by Simon (1884).
Linyphia proserpina Simon, 1872 — Simon (1872): p. 475, Pl. 16, Figs. 6–7 View FIGURE 6 View FIGURE 7 (descr. ♂); synonymised by Holm (1944).
L. pusiola Fickert, 1875 — Fickert (1875): p. 20, Fig. 3 View FIGURE 3 (descr. ♂); synonymised by Rozwałka (2010).
Bathyphantes charpentieri Lebert, 1877 — Lebert (1877): p. 163, Pl. 4–5, Figs. 29–35 View FIGURE 29 View FIGURE 30 View FIGURE 31 View FIGURE 32 View FIGURE 33 View FIGURE 34 View FIGURE 35 (descr. ♂ ♀); new synonymy.
L. thorellii Herman, 1879 —Herman (1879): p. 69, 345, Plate VII, Fig. 153 (descr. ♂ ♀); synonymised by Fage (1931).
B. convexus — Simon (1884): p. 351; synonymy.
Porrhomma proserpina — Simon (1884): p. 360, Fig. 134 ( ♂).
P. scopiferum Simon, 1884 — Simon (1884): p. 361, Fig. 136–137 (descr. ♂ ♀); synonymised by Fage (1931).
P. diabolicum Simon, 1884 — Simon (1884): p. 362 (descr. ♂ ♀); synonymised by Fage (1931).
P. norvegicum Strand, 1901 View in CoL — Strand (1901), p. 25, Figs. 5a–c View FIGURE 5 (descr. ♂ ♀); examined and synonymised by Holm (1944).
P. proserpina View in CoL — Fage (1931): p. 147, 158, Figs. XIV–XV ( ♂ ♀); synonymy.
P. pygmaeum forma proserpina — Miller & Kratochvíl (1940): p. 167, Figs. 1–2 View FIGURE 1 View FIGURE 2 ( ♂ ♀).
P. ocella Chamberlin & Ivie, 1943 View in CoL — Chamberlin & Ivie (1943): p. 4, Figs. 1–2 View FIGURE 1 View FIGURE 2 (descr. ♂, not ♀); new synonymy.
Willibaldia (?) sodonta Chamberlin, 1949 View in CoL — Chamberlin (1949): p. 559, Fig. 146 (descr. ♀); transferred by Thaler (1968); new synonymy.
P. convexum View in CoL — Holm (1944): 130, 133; synonymy.
P. pygmaeum convexum View in CoL — Thaler (1968): 367, 369, Figs. 2a View FIGURE 2 , 4d–e View FIGURE 4 , 5a–b View FIGURE 5 ( ♂ ♀).
P. sodonta — Thaler (1968): p. 385.
P. convexum — Miller (1971): p. 238, pl. XLVII, Figs. 12–13 View FIGURE 12 View FIGURE 13 (♂ ♀).
sub P. convexum —Loksa (1981): p. 335, Figs. 12–14 View FIGURE 12 View FIGURE 13 View FIGURE 14 ( ♂ ♀); misidentification, it refers to P. microps .
Material examined. Holotype Ƌ, labelled as Linella ocella Chamberlin & Ivie , UTAH: City Creek Can. ( Rotary
Park), 111.40 nw. September 16, 1942, Wilton Ivie collector. Paratypes 1 Ƌ 3 ♀, labelled as Linella ocella Chamberlin & Ivie , UTAH: Smith and Morehouse Canyon , 111.40 nw. , October 7, 1932, Wilton Ivie collector; only a male, females are Anacornia microps ( Chamberlin & Ivie, 1933) , misidentification (AMNH).
Other material examined. CZECHIA: Pec pod Sněžkou, Krkonoše Mts. , Úpská Jáma corrie, 4–19 Aug 1986, 1 Ƌ, leg. J. Vaněk. Teplice nad Metují, Adršpašské Skály rocks, 13 Jul 1986, 1 Ƌ, leg. V. Růžička. Teplice nad Bečvou, Hranická Propast abyss, 16 Mar 2003, 3 ♀; Adamov , Býčí Skála Cave , 10 Nov 2006, 5 ♂ 3 ♀, leg. R. Mlejnek ( IECA) . SLOVAKIA, Slovak Karst, Domica Cave, 8 Mar 2001, 2 Ƌ 1 ♀, 11 Aug 2011, 1 ♀, leg. Ľ. Kováč & P. Ľuptáčik ( WSM) . BULGARIA: Mladezhko (distr. Malko Tarnovo), Kaleto Cave , 10 Aug 1985, 1 ♂ 3 ♀; Jglika (distr. Jambol), Leleshka Dupka Cave, 8 Aug 1970, 1 ♂ 2 ♀; Stubel (distr. Montana), Mecha Dupka Cave, 11 Jun 1972, 1 ♂ 2 ♀; Lipnitsa (distr. Sifia), Kozarnika Cave, 14 Oct 1973, 1 ♂ 2 ♀, leg. et coll. C. Delthsev . ROMANIA. Damiş env., Roşsia, Ciur Ponor Cave, 46°51'N, 22°24'E, 24 Jun 2011, 1 Ƌ, leg. M. Boukal (CJD) GoogleMaps . NORWAY: Svartisen National Park , Aug 1962, 2 Ƌ 1 ♀ ( AMNH) . SWEDEN: Kaisepakte, Torne River , 1 Ƌ, leg. A. Holm ( AMNH) . RUSSIA: SW Tuva, Western Sayan Mts., Bulba River , 52°47'N, 93°18'E, 1230 m, 21 Jun 1995, 1 Ƌ 2 ♀, leg. Yu. M GoogleMaps . Marusik (CYuM). USA: Colorado, Estes Park , 40°N, 105°W, 27 August 1961, 1 ♀, leg. W.J. Gertsch & W . Ivie. Illinois, Monroe County, Dry Run Cave , 38°N, 90°W, 26 Jun 1965, 1 ♀, leg. S. Peck . Kentucky, Woodfort County, Swope Cave , 3 Jul 1957, 1 ♀, leg. C. Krekeler & J. Rittmann . Maryland, Baltimore , 39.18°N, 76.38°W, 12 May 1966, 4 ♀, leg. B. Kamfchick GoogleMaps . Oregon, Hebo , 45.13°N, 123.53°W, 3 Apr 1948, 1 Ƌ 1 ♀, leg. V. Roth GoogleMaps . Tennessee, Claiborne County, Coonsies Creek, Creek Cave , 2 Aug 1974, 1 Ƌ, leg. J.R. Holsinger & D . C. Culver. Utah, City Creek Canyon ( Rotary Park ), 16 Sep 1942, 1 Ƌ, leg. W. Ivie . Vermont, Rutland County, Norris Cave (¾ mi. S Danby), 23 Apr 1967, 1 ♀, leg. S. Peck . Washington, Waterville , 47°N, 120°W, 28 Aug 1959, 2 Ƌ 1 ♀, leg. W.J. Gertsch & V. Roth ( AMNH) . CANADA: Alberta, Waterton Lakes National Park , 10–13 Jul 1973, 1 Ƌ, leg. J.H. Redner & C . Starr. British Columbia, Pine Pass, between Chetwynd and McLeod Lake , 11 Jul 1981, 1 Ƌ 2 ♀, leg. C.D. Dondale ( CNC); Goldstream Park , 48°N, 123°W, 23 Apr 1951, 1 ♀, leg. R. Guppy ( AMNH).
Diagnosis. A large group of species is characterised by embolus of middle length and S-shaped ascending parts of copulatory ducts: P. borgesi , P. cambridgei , P. convexum , P. errans , P. nekolai , P. oblitum , P. pygmaeum and P. rosenhaueri . P. convexum can be distinguished from all these species by the following combination of characters: CW> 0.69 mm, Mt I/CW = 0.91–1.06. A pair of almost parallel spots, representing translucent copulatory ducts, is visible on the epigynum ( Fig. 19C View FIGURE 19 ).
Description. ♀ (from Teplice nad Bečvou, Czechia, 16 Mar 2003). Carapace brown, 0.81 mm wide, eyes normal, PME–PME = 1.0 ( Fig. 19A View FIGURE 19 ). Abdomen greyish-yellow. Fe I–II with one dorsal spine, Fe I with one prolateral spine. Ti I with one prolateral spine, Ti I–II with one retrolateral spine. Tm Mt I = 0.46, Mt I/CW = 1.01.
A pair of almost parallel spots, representing translucent copulatory ducts, is visible on the epigynum ( Fig. 19C View FIGURE 19 ). Ascending parts of the ducts are S-shaped. Spermathecae are formed behind the ascending part of the ducts ( Figs. 19D–F View FIGURE 19 ).
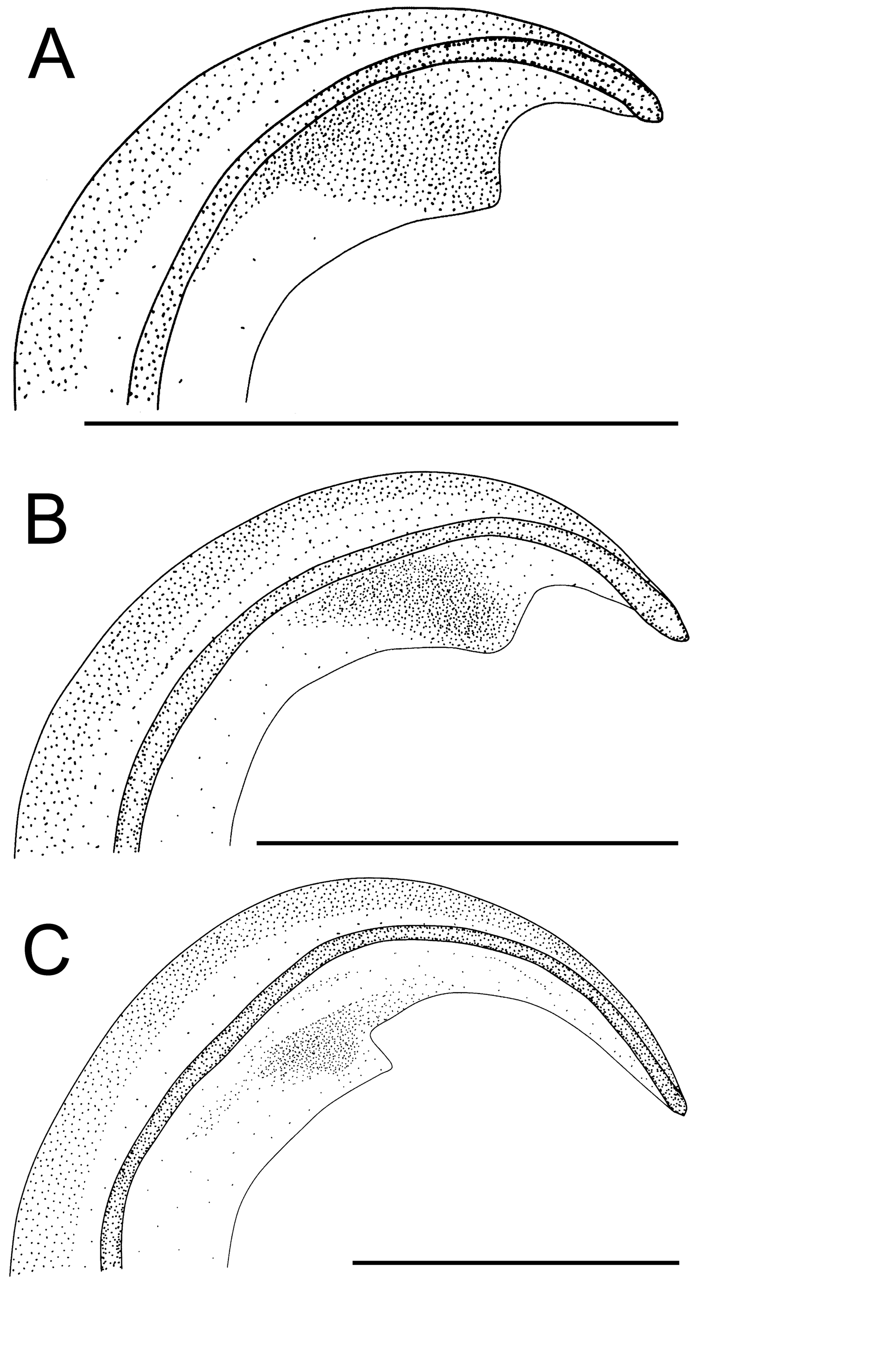
Ƌ (from Pec pod Sněžkou, Úpská Jáma corrie, Czechia, 4–19 Aug 1986). Embolus of middle length with a narrow velum. AP has the form of a bird head ( Fig. 19B View FIGURE 19 ).
Variation. Ƌ ♀. Carapace 0.69–0.92 mm wide. Tm Mt I = 0.40–0.48, Mt I/CW = 0.91–1.06 (n = 13).
Comments. Simon (1884: p. 364) synonymised B. charpentieri with P. errans . Roewer (1942: p. 600) synonymised B. charpentieri with P. microphthalmum , and this synonymy was accepted in the World Spider Catalog (2017). However, Lebert (1877) presented a new species on precise line drawings. The embolus, depicted on Figs. 31 View FIGURE 31 and 32 View FIGURE 32 , is lacking a prominent velum; the epigynum on Fig. 33 View FIGURE 33 is lacking a square form, which is typical for P. microphthalmum . I must remove Bathyphantes charpentieri from the synonymy of P. microphthalmum . The material was collected in subterranean spaces in a salt mine and is characterised by depigmentation and width of male cephalothorax of 0.8–0.96 mm (out of the range of P. microphthalmum ). Also, the epigynum is characteristic for P. convexum . Lessert (1910: p. 619), Fage (1931: p. 158) and Thaler (1968: p. 384) also supported this synonymy.
The male specimen of Linyphia pusiola Fickert, 1875 was collected under a stone in the Giant Mountains ( Czech Republic) at an altitude of 1400 m. In this subalpine belt, only P. egeria , P. convexum and P. microphthalmum occur ( Buchar & Růžička 2002). According to the figure of the male palp, I support synonymy with P. convexum proposed by Rozwałka (2010).
Holotype of Porrhomma ocella was compared with specimens of Porrhomma convexum and is identical in all aspects, mainly in the structure of the male palp. The females associated with the holotype male are in fact Anacornia microps Chamberlin & Ivie, 1933 . This was noted by Crawford (1988) and repeated by Buckle et al. (2001). Examined females significantly differ from P. convexum: The overall colouration is orange-brown and the PME are more than 2.5 diameters apart. All characteristics and the form of the epigynum of allotype and paratype females agree with the description and illustrations of Anacornia microps (see Chamberlin & Ivie 1933; Chamberlin 1949).
Type material of Porrhomma sodonta was not available. The figure of the epigynum, together with the carapace width of 0.87 mm, indicates the conspecificity with P. convexum . The species was described according to one female, collected in White R. Camp, Rainier Park, July 6, 1938 ( Chamberlin 1949). In the material examined, one female of P. convexum with this designation was present: “ Porrhomma ? ♀, Washington: White Riv. Camp, Rainier Park, 46°54'N, 121°37'W, July 6, 1938, Wilton Ivie Collector.” It is probably the lost holotype.
Loksa (1981, p. 335, Figs. 12–14 View FIGURE 12 View FIGURE 13 View FIGURE 14 ) should be striked out from taxonomic references for P. convexum . The specimens extracted from forest litter are clearly P. microps , according to the broad side loops of copulatory ducts, long embolus with a broad velum and long anterior process of the embolic plate.
Specimens from Canada and Austria show high correspondence in all characteristics considered ( Breuss 2009).
Ecology. Porrhomma convexum colonises habitats with constantly high humidity and low temperature, such as entrances of caves and mines, river banks and rock slopes. It has also been found in caves at a depth of 80 m ( Růžička et al. 2013).
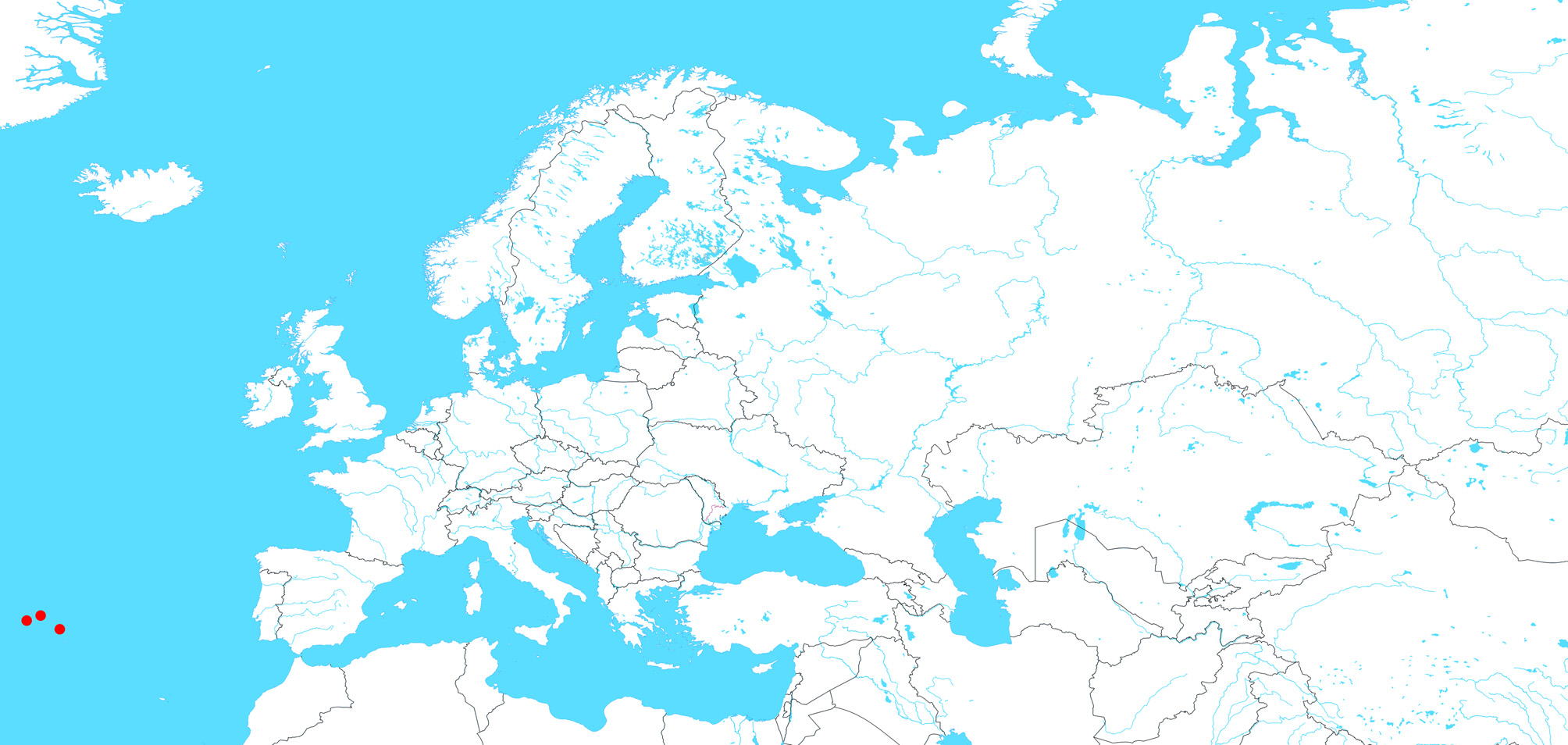
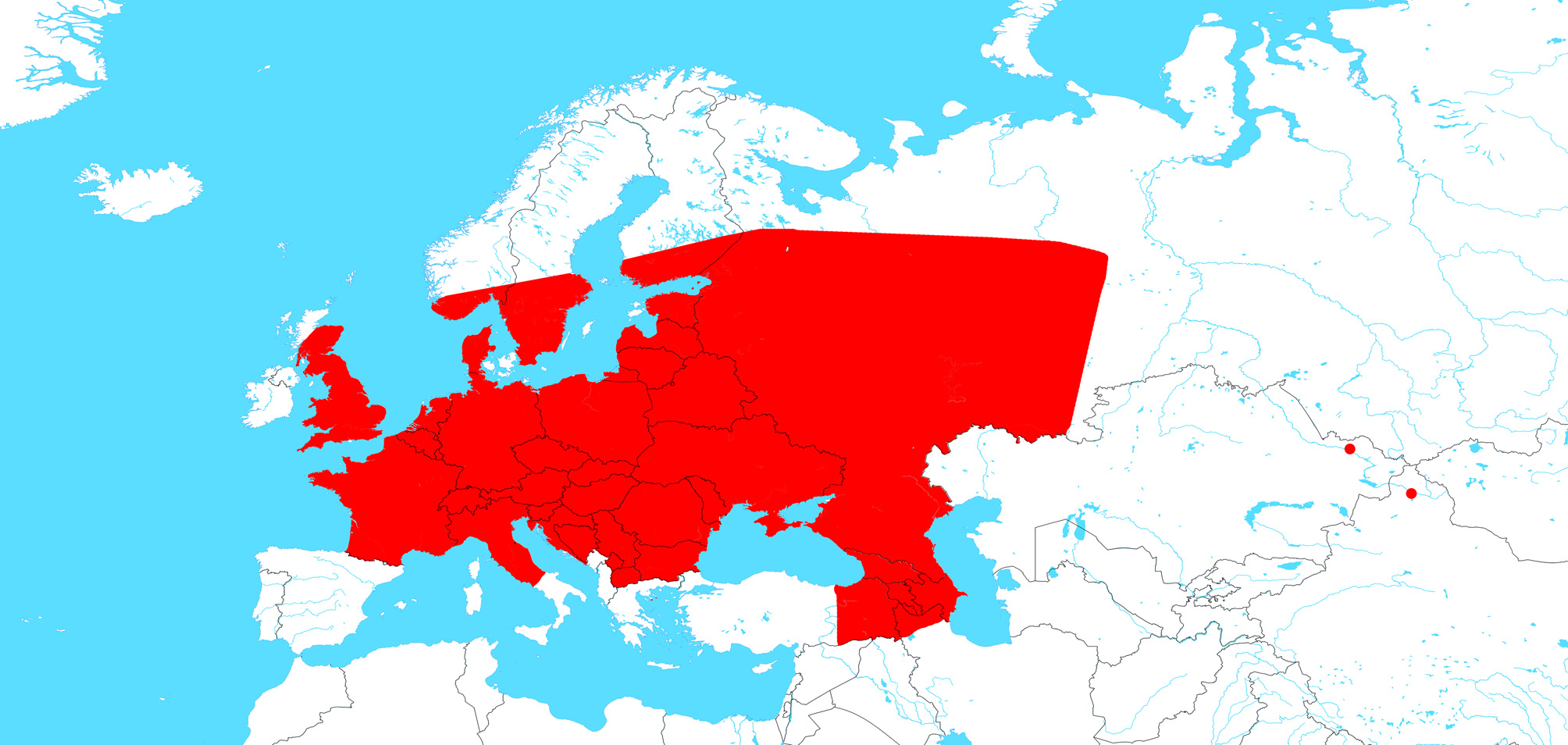
Global distribution. Eurasia after Helsdingen (2017) and Mikhailov (2013), Greenland after Marusik et al. (2006), North America after Breuss (2009) and this article. See Fig. 20 View FIGURE 20 .












No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Porrhomma convexum ( Westring, 1851 )
| Růžička, Vlastimil 2018 |
P. convexum
| Holm (1944) : 130 |
P. pygmaeum convexum
| Thaler (1968) : 367 |