
Pista corrientis McIntosh, 1885
|
publication ID |
https://doi.org/10.5281/zenodo.278414 |
|
DOI |
https://doi.org/10.5281/zenodo.5633380 |
|
persistent identifier |
https://treatment.plazi.org/id/03C4D91A-D505-FFA4-17FF-F9D1FDFD94FD |
|
treatment provided by |
Plazi |
|
scientific name |
Pista corrientis McIntosh, 1885 |
| status |
|
Pista corrientis McIntosh, 1885 View in CoL
Figures 1–2 View FIGURE 1 View FIGURE 2
Pista corrientis McIntosh 1885: 457 View in CoL –458, Pl. XLVIII, fig. 11; Pl. XXVII A, Fig. 35.
Not Pista corrientis: Hessle 1917: 158 View in CoL –159, Taf. II, Figs. 2–3 View FIGURE 2 View FIGURE 3 . Hartman 1966: 97 –99, Pl. XXXIII, Figs. 4–9 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 . Blankensteyn 1988: 51 –54, fig. 10. Nogueira 2000: p. 180–183, Figs. 42–43. Alves 2008: 70 –74, Figs. 21B, 22–23.
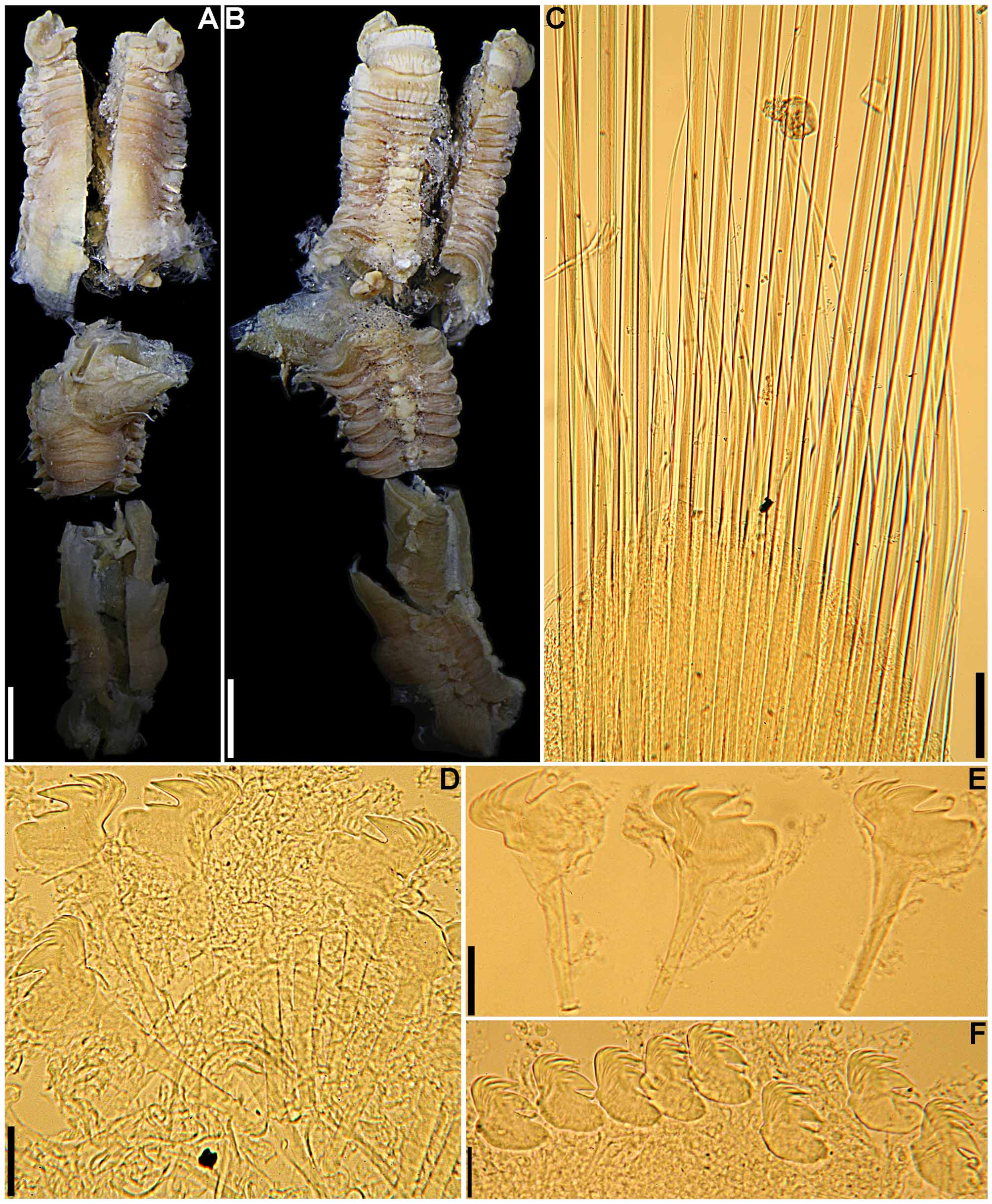
Material examined. Holotype ( BMNH 1885.12.1.348): complete specimen in poor state of preservation (see below), 27 mm long, 1.5 mm wide, with 93 segments; coll. Argentina, off mouth of Rio de la Plata, 37o17’S 53o52’W, ~ 1100 m (600 fm), sea bottom, green sand, Challenger Expedition, 14 Feb 1876. Slides: neurochaetae from segments 9, 19, 23 and 83.
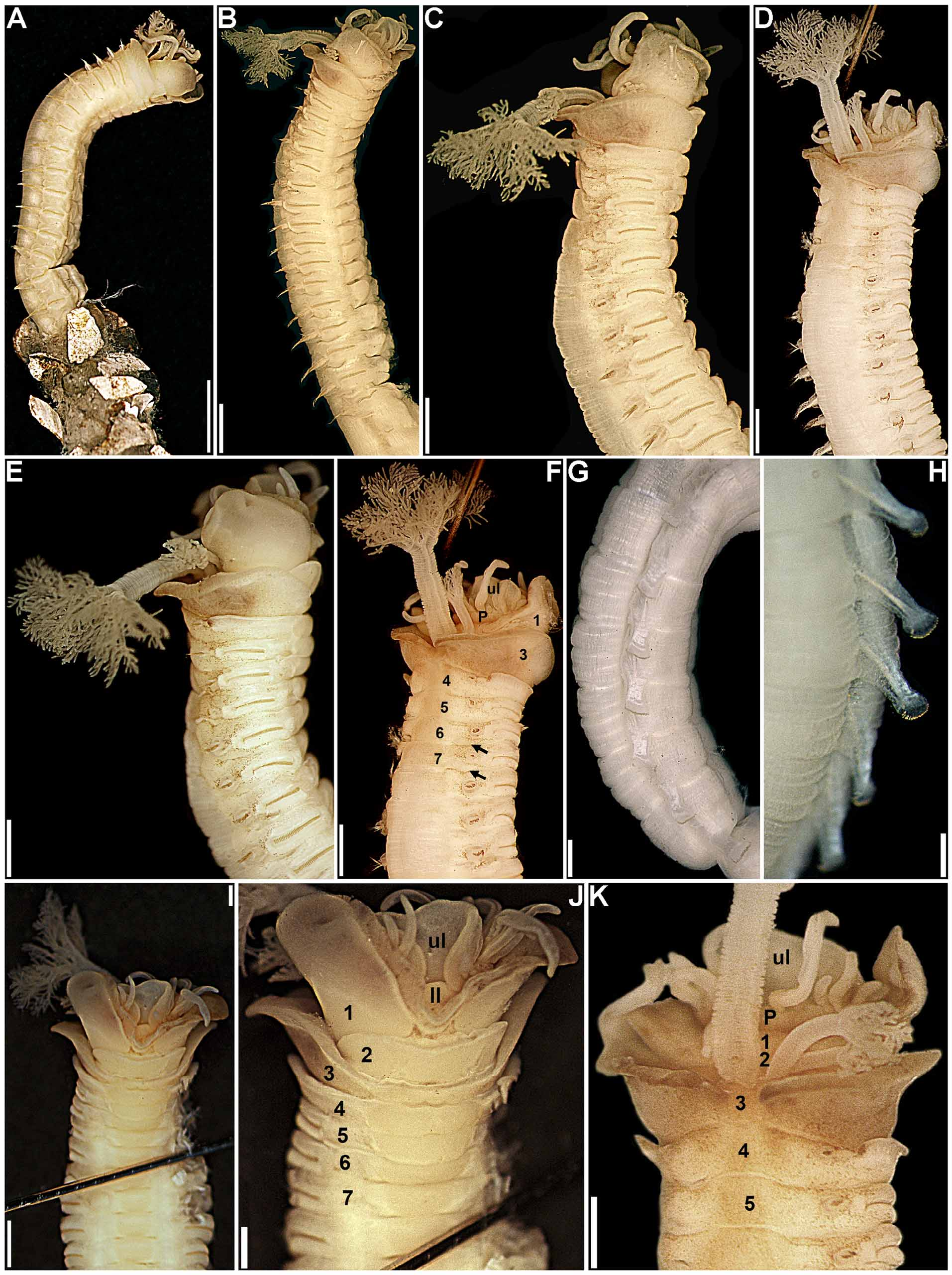
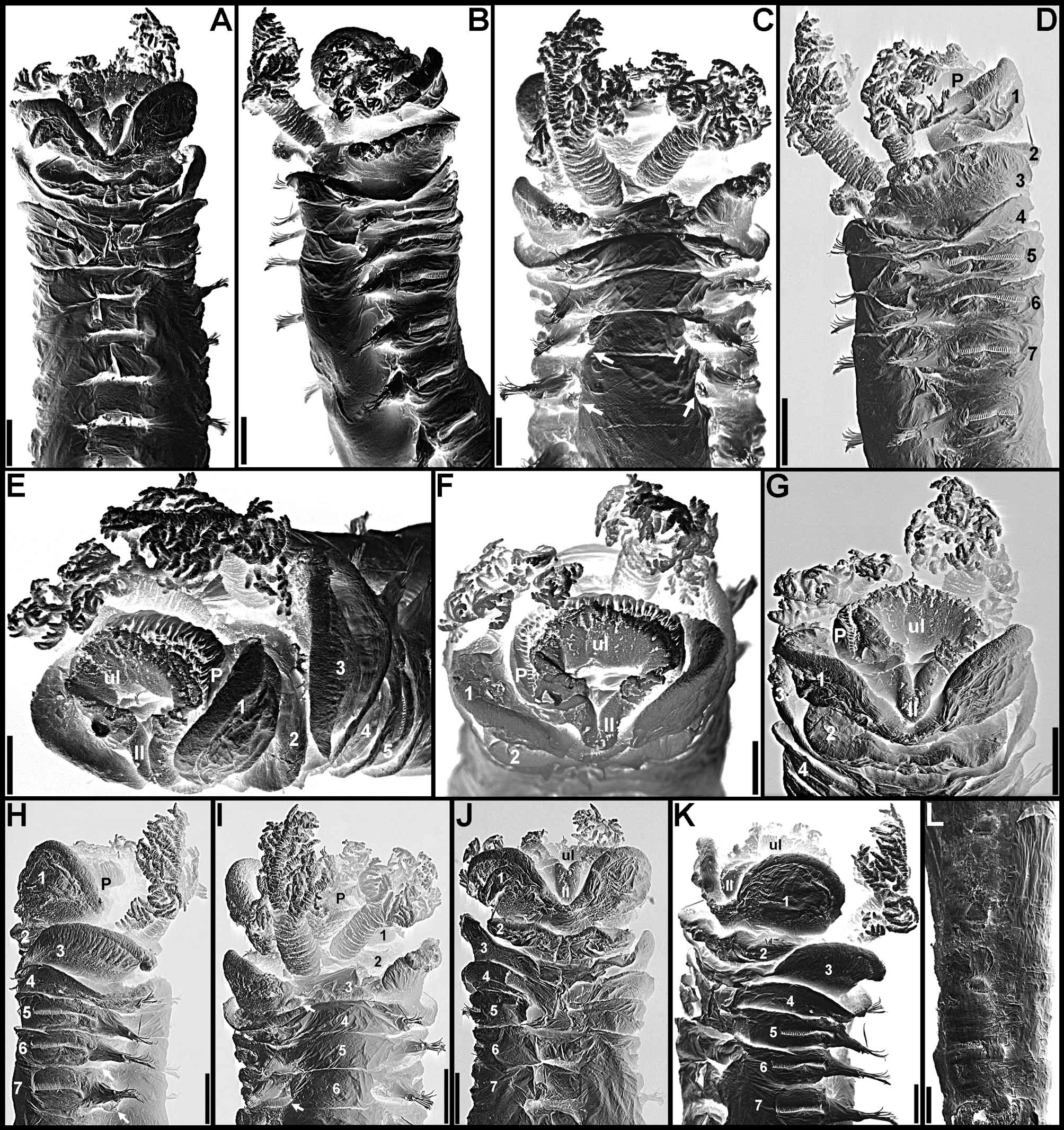
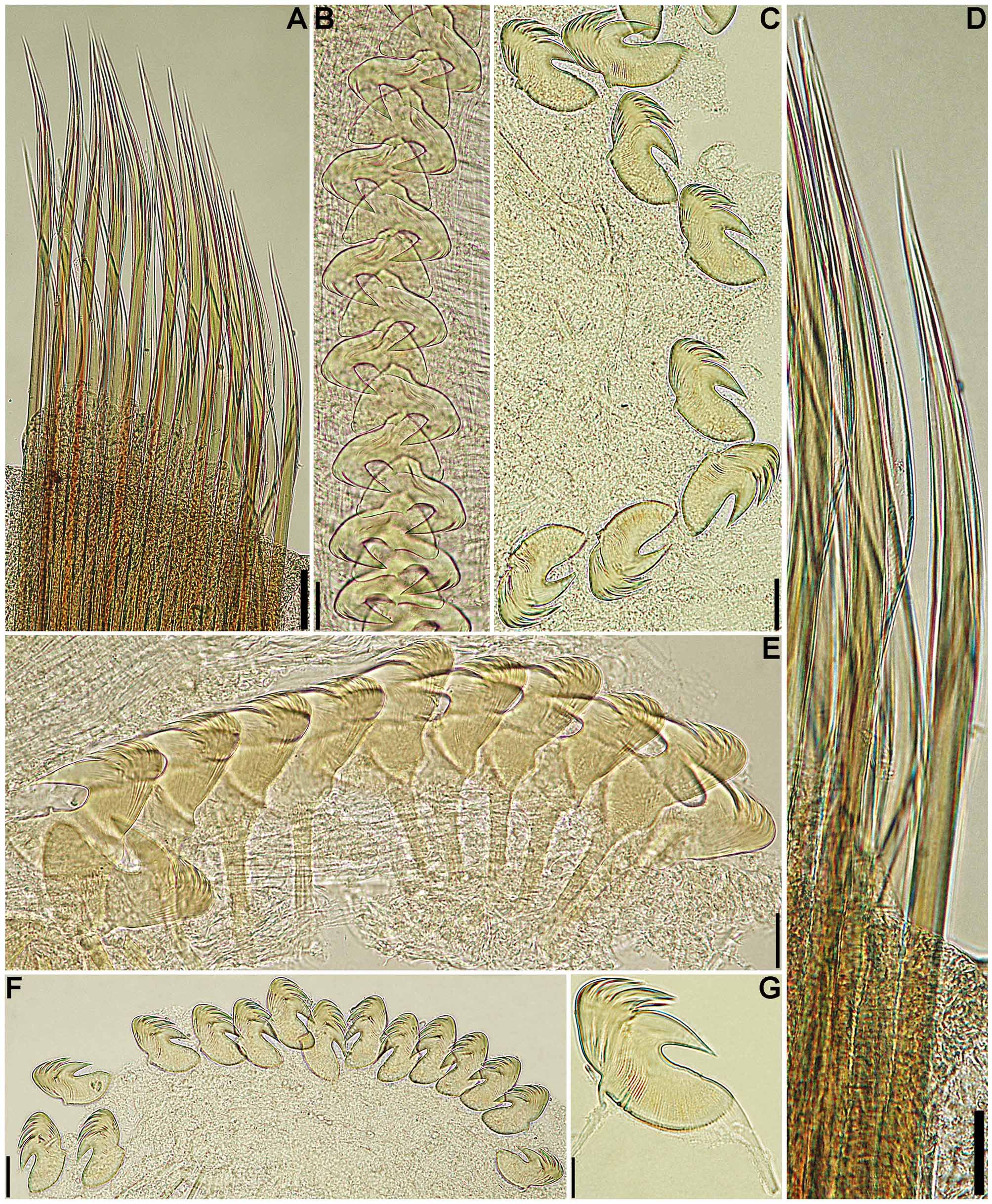
Description. Holotype in very poor condition, all buccal tentacles missing and anterior body opened by two longitudinal cuts, one on each side, extending from segment 1 to last segment with biramous parapodia ( Fig. 1 View FIGURE 1 B–C, F–G, I–K), almost torn in two pieces at beginning of region with neuropodia only, dorsal body wall missing between segments 25 and 30. Preserved body light brown. Body not dorsally inflated; anterior segments compact, about same length, progressively longer on segments 8–18, elongation more evident from segment 11 ( Fig. 1 View FIGURE 1 A–C, F–L). Ventral shields on segments 1–20, first shield crenulated, following shields smooth, all rectangular; widest shield on segment 5, progressively narrower until segment 10, then about same width until segment 15; shortest shield on segment 3, almost inconspicuous, progressively longer until segment 15, then progressively shorter and less conspicuous until last shield ( Fig. 1 View FIGURE 1 A–C, G–I, K–L); after segment 20 shields replaced by mid-ventral groove extending posteriorly. Prostomium at base of upper lip; distal part forming shelf-like process from which buccal tentacles originate; all buccal tentacles missing; no eyespots visible on basal part of prostomium ( Fig. 1 View FIGURE 1 B–C, F–G, I–K). Peristomium restricted to lips; upper lip short, longer than wide; lower lip short, cushion-like, partially covered by lobes on segment 1 ( Fig. 1 View FIGURE 1 A–C, F–L). Segment 1 narrow, with one pair of large, distally rounded lobes originating dorso-laterally, extending anteriorly and ventrally, terminating below level of upper lip, connected to each other by lower mid-ventral membrane partially exposing lower lip ( Fig. 1 View FIGURE 1 A–C, F–L). Segment 2 dorsally and laterally short, longer ventrally, with one pair of short and rounded ventro-lateral lobes, connected to each other by low membrane across ventrum ( Fig. 1 View FIGURE 1 A–C, F–L). Segment 3 with one pair of large, almost triangular, distally rounded lobes originating at level of notopodia on segment 4 and terminating at level of ventral edge of neuropodia, well separated from ventral shields; anterior margin of segment 3 forming mid-dorsal crest ( Fig. 1 View FIGURE 1 A–C, F–L). Segment 4 with short lateral lobes, as thin flaps, connected to each other by dorsal crest on anterior margin of segment, more conspicuous than dorsal crest on segment 3 ( Fig. 1 View FIGURE 1 A–C, F–L); segments 5–6 with short ventro-lateral lobes as low flaps between ventral shields and neuropodia. Branchiae missing, scars evident on segments 2–3, anterior pair dorsally aligned to second pair. Seventeen pairs of notopodia, on segments 4–20, all similar in size; notopodia of segments 4–7 inserted progressively more laterally ( Fig. 1 View FIGURE 1 B–C, F–G, I–K), then vertically aligned. Distal portions of all notochaetae broken off. Neuropodia starting from segment 5, as low rectangular ridges slightly raised from surface of body until segment 20 ( Fig. 1 View FIGURE 1 A–C, F–L), as short pinnules on anterior region with neuropodia only ( Fig. 1 View FIGURE 1 D), longer on posterior body segments ( Fig. 1 View FIGURE 1 E); neuropodial pinnules situated laterally on body, well separated from mid-ventral groove, internal neuropodial shafts present. Long-handled uncini on segments 5–20, with thin handle originating from heel, short handled thereafter; in region with biramous parapodia, uncini with strong dorsal button at mid-length of area between base of main fang and tip of prow, distally rounded prow, and crest with 3 rows of secondary teeth ( Fig. 2 View FIGURE 2 A, C); uncini arranged in completely intercalated double rows from segment 11 until segment on which notopodia terminate; from segment 21, uncini similar to those from region with biramous parapodia, but short-handled and with smaller dorsal button ( Fig. 2 View FIGURE 2 B, D). One pair of nephridial papillae on segment 3, inserted dorsal to bases of branchiae ( Fig. 1 View FIGURE 1 F, J); genital papillae situated posteriorly and dorsally to notopodia on segments 6–7 ( Fig. 1 View FIGURE 1 B–C, F–G, I–K). Pygidium smooth ( Fig. 1 View FIGURE 1 E).
Remarks. The original description of P. corrientis does not mention most of the characters currently considered important for the taxonomy of the genus. Further descriptions of this taxon were provided by Hessle (1917) and Hartman (1966); however, in both cases the descriptions provided do not match the characters of the holotype and cannot belong to this species.
The text provided by Hessle (1917) matches the holotype very closely, but he did not mention several characters, including the segments on which long-handled uncini occur. However, the drawings provided by Hessle show remarkably different morphology of the lobes on anterior segments, especially those on segment 1, which, according to Hessle, are almost rectangular, distally straight, while those of the holotype of Pista corrientis are almost triangular and distally rounded.
Hartman (1966) described P. corrientis as having large, lateral lobes projecting forward on segment 1, although from her illustration ( Hartman 1966: 98–99, Pl. XXXIII, fig. 4, copied from Ehlers, 1913) it is not certain if they terminate below the level of the upper lip, as in the holotype, or if they extend beyond it. Hartman said longhandled uncini are present on anterior segments and then change to short-handled, but she did not say on which segment that occurs. Hartman, however, included illustrations of uncini from a non-specified mid-thoracic segment and also from segment 19 and both are short-handled ( Hartman 1966: 98–99, Pl. XXXIII, figs. 5 [copied from Ehlers, 1913] and 8 [copied from Monro, 1930]). Considering that the holotype of P. corrientis has long-handled uncini on segments 5–20, the specimens studied by Hartman cannot belong to this taxon. We assume that if Hartman included those figures in her paper that is because she examined her material and agreed with those illustrations, which leads us to conclude that neither Hartman (1966), Monro (1930) or Ehlers (1913) had true P. corrientis .
Probably because of the erroneous description provided by Hartman (1966), P. corrientis was later reported from the Brazilian coast ( Nonato 1973; Lana 1981; Blankensteyn 1988; Sovierzoski 1991; Santos et al. 1994; Nogueira 2000; Rizzo & Amaral 2001; Amaral et al. 2003; Alves 2008). The Brazilian material is herein described as P. nonatoi spec. nov. (see below).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Terebellinae |
|
Genus |
Pista corrientis McIntosh, 1885
| Nogueira, João Miguel De Matos, Harris, Leslie, Hutchings, Pat & Fukuda, Marcelo Veronesi 2011 |
Pista corrientis :
| Alves 2008: 70 |
| Blankensteyn 1988: 51 |
| Hartman 1966: 97 |
| Hessle 1917: 158 |
Pista corrientis
| McIntosh 1885: 457 |