
Trichogramma marandobai Brun, Moraes & Soares, 1986
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4021.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:16BED02E-5E22-4EE6-B050-2BFE3B61374C |
|
DOI |
https://doi.org/10.5281/zenodo.5697872 |
|
persistent identifier |
https://treatment.plazi.org/id/03C2EC3D-FFC3-585E-FF24-FEE4FD24F816 |
|
treatment provided by |
Plazi |
|
scientific name |
Trichogramma marandobai Brun, Moraes & Soares, 1986 |
| status |
|
Trichogramma marandobai Brun, Moraes & Soares, 1986 View in CoL
Brun, Moraes & Soares 1986: 807; Zucchi & Monteiro 1997: 54 (catalogue); Querino & Zucchi 2005: 50, Zucchi et al. 2010: 233; Querino & Zucchi 2011: 58 (catalogue).
Trichogramma demoraesi: Zucchi & Monteiro 1997: 53 View in CoL (catalogue); Querino 2002: 72; Querino & Zucchi 2005: 51; Ronchi- Teles & Querino 2005: 515; Zucchi et al. 2010: 231 (catalogue); Querino & Zucchi 2011: 38 (catalogue); Zucchi 1985:43 (thesis).
Diagnosis. Trichogramma marandobai is similar to T. demoraesi , but differs in that the flagelliform setae are shorter in T. marandobai than T. demoraesi . In T. marandobai the intervolsellar process is robust, truncated or pointed at the apex; the genital capsule is long, with a typical dorsal lamina with a wide basis and a posterior extension with a pointed apex, not exceeding the apex of the volsella, and the posterior extension wider at the base.
Antenna ( Fig. 1 View FIGURE 1 A): flagellum length (0.15 ± 0.003); ratio of length to width of flagellum (4.01 ± 0.06); flagellum length to hind tibial length (0.90 ± 0.02); flagelliform setae short (0.06 ± 0.001) ( Fig. 1 View FIGURE 1 B); longest flagelliform setae length to greatest flagellum width (1.55 ± 0.03); basiconic sensilla with formula 1-2-2-0(1)-1-1 ( Fig. 1 View FIGURE 1 C); three placoid sensilla, basal sensillum (0.05 ± 0.001). Fore wing: width (0.27 ± 0.01); width to length ratio (0.51±0.01); fringe setae length (0.05 ± 0.001); fringe setae length to length of hind tibia (0.27 ± 0.01); fringe setae length to width of fore wing (0.18 ± 0.01); 8–31 setae between 4th and 5th setal tracks. Hind wing: anterior track with 2 to 5 setae, posterior track with 6 to 11 setae, reaching midpoint of middle track.
Mesoscutum ( Fig. 1 View FIGURE 1 D): lengths of anterior and posterior pair similar. Scutellum ( Fig. 1 View FIGURE 1 E): ratio of anterior to posterior pair (0.24 ± 0.01).
Male genitalia ( Figs. 2 View FIGURE 2 A–C): genital capsule longer (0.14 ± 0.002) than wide (0.06 ± 0.001); ratio of width to length of genital capsule (0.43 ± 0.01); apical distance to genital capsule length (0.27 ± 0.02); apical width to genital capsule length (0.27 ± 0.02); dorsal lamina length to genital capsule length (0.37 ± 0.01); dorsal lamina length to dorsal lamina width (1.34 ± 0.03); dorsal aperture length to dorsal lamina length (1.54 ± 0.03); posterior extension of dorsal lamina (0.04 ± 0.001); dorsal lamina medium (0.05 ± 0.001), narrowing from apex to base; posterior extension of dorsal lamina length to dorsal lamina length (0.86 ± 0.01); dorsal ridge absent; ventral ridge medium (0.05 ± 0.001), reaching midpoint of genital capsule; ventral ridge length to basal distance (0.51 ± 0.03); basal distance to genital capsule length (0.73 ± 0.02); intervolsellar process long (0.01 ± 0.00), distinct, and pointed or truncated apex ( Fig. 6 View FIGURE 6 ), not reaching apex of volsella; ventral processes near base of intervolsellar process (0.01 ± 0.00).
Material examined. BRAZIL, São Paulo: Atibaia (Instituto Biológico), 5 ♂; Piracicaba, 1.vii.1999, 15 ♂ (as T. demoraesi ) (R. B. Querino); Minas Gerais: Janaúba, 9.iii.2010 15 ♂ (J. M. Vieira) ( ESALQ).
Type material. Apparently lost; not deposited in the collection of Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais (Pedro M. Linardi, pers. comm.). Inquiries from other sites for location of type material, provided by George W.G. Moraes, were unsuccessful.
Geographical distribution. Brazil—Amazonas ( Ronchi-Teles & Querino 2005), Mato Grosso do Sul ( Oliveira et al. 2010), Minas Gerais ( Brun et al. 1986; Vieira 2011), Paraná ( Querino & Zucchi 2008, collection), São Paulo ( Querino & Zucchi 2008, collection).
Multivariate morphometry. Based on the analysis of canonical variables, specimens of T. marandobai comprised three groups, according to collection sites: (1) Janaúba, MG, with specimens with IT and IP processes, (2) Piracicaba, SP; and (3) Atibaia, SP ( Fig. 3 View FIGURE 3 ). The first component explained 68% of the total variance in the covariance matrix. The variables placoid sensilla length, antenna scape length and dorsal lamina length showed the highest absolute values (0.98, 0.92 and 0.92, respectively).
These variables separated the specimens from the three collection municipalities (Janaúba, Piracicaba and Atibaia). However, the variables grouped the specimens from Janaúba with variations in the intervolsellar process, confirming the presence of intraspecific variations, and therefore, these specimens were classified as co-specific. The second component had lower explanatory power (21%), represented by apex width, length and aedeagus length of the posterior portion of the dorsal lamina, which showed the highest absolute values (1.1, 0.90 and 0.89, respectively).
Geometric morphometry. The analysis of canonical variables VC1 and VC2 generated by the positional coordinates of the Cartesian plane of 15 anatomical landmarks of the male genitalia of T. marandobai from the three municipalities (Janaúba, Piracicaba and Atibaia) revealed similarities among the populations ( Fig. 4 View FIGURE 4 ). These results were similar to those obtained with the multivariate morphometry, even for specimens with variations in the intervolsellar process collected in Janaúba. The Wilks’ Lambda test (P <0.0001) showed that the canonical axes were significant.
In the analysis of relative deformation, deformation diagrams ( Fig. 5 View FIGURE 5 ) showed that individuals located at the lower end of VC1 ( Fig. 4 View FIGURE 4 ) had a wider genital capsule than the others.
Values of the Euclidean distances calculated from the shape component of the genital capsule of T. marandobai from Janaúba (IT or IP processes), Piracicaba and Atibaia were used in the cluster analysis ( UPGMA). According to this analysis, individuals from Janaúba with IT and IP processes were grouped in the same clade, and closest to those from Atibaia, which were in turn closer to those from Piracicaba ( Fig. 6 View FIGURE 6 ).
Crosses. Individuals were consistent in bidirectional crosses, as they originated individuals of both sexes, that is, the females were fertilized in heterogamic crosses ( Table 2 View TABLE 2 ). The individual virgin females originated fertile offspring by parthenogenesis, confirming the reproduction means of the species. Therefore, crosses performed with morphotypes of T. marandobai helped to determine the identity and definition of the species.
Offspring of isofemale strains of T. marandobai from homogamic crosses showed variations in the intervolsellar process, that is, individuals had a pointed or truncated intervolsellar process ( Fig. 8 View FIGURE 8 ). Therefore, the crosses established that these apex shapes of the intervolsellar process are intraspecific variations.
Molecular analysis. The ITS2 amplification reactions of strains produced an amplicon of approximately 550 bp, of which 503 bp were sequenced and aligned for individuals with IT and IP processes. The ITS2 amplicons for the two morphotypes of T. marandobai were identical, confirming the morphometric data, which indicate that the morphological variation in male genitalia of T. marandobai is an intraspecific variation.
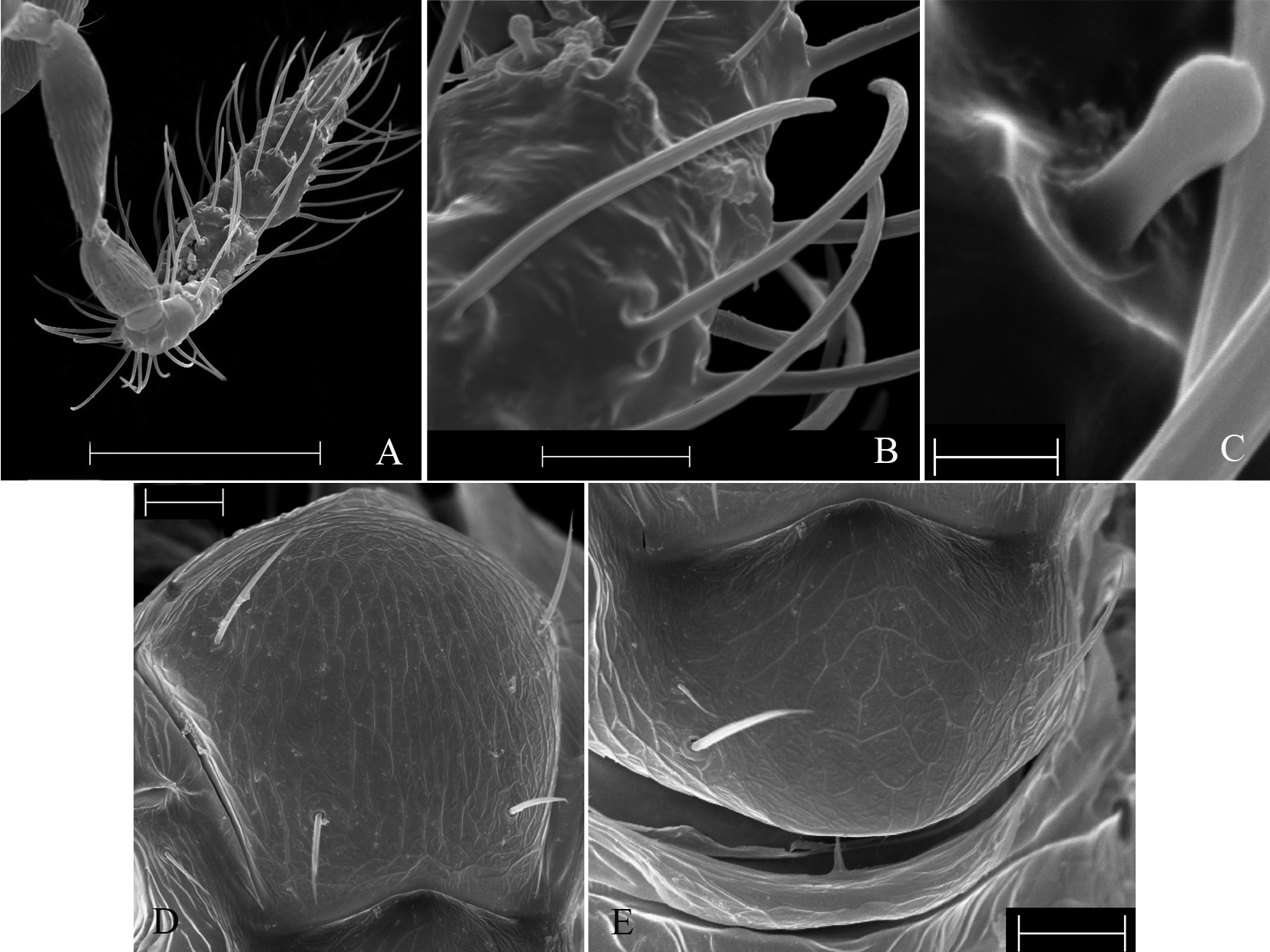
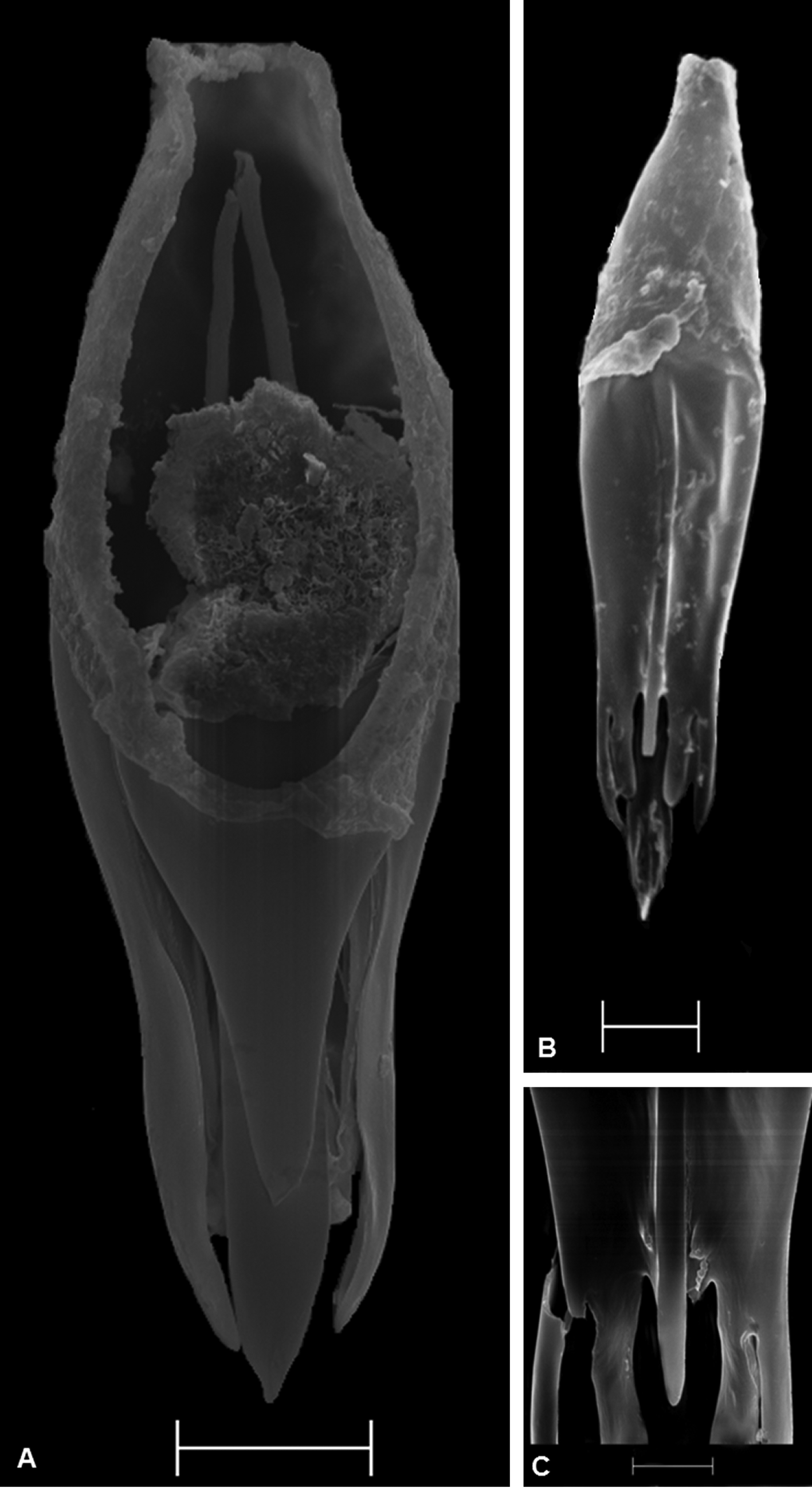
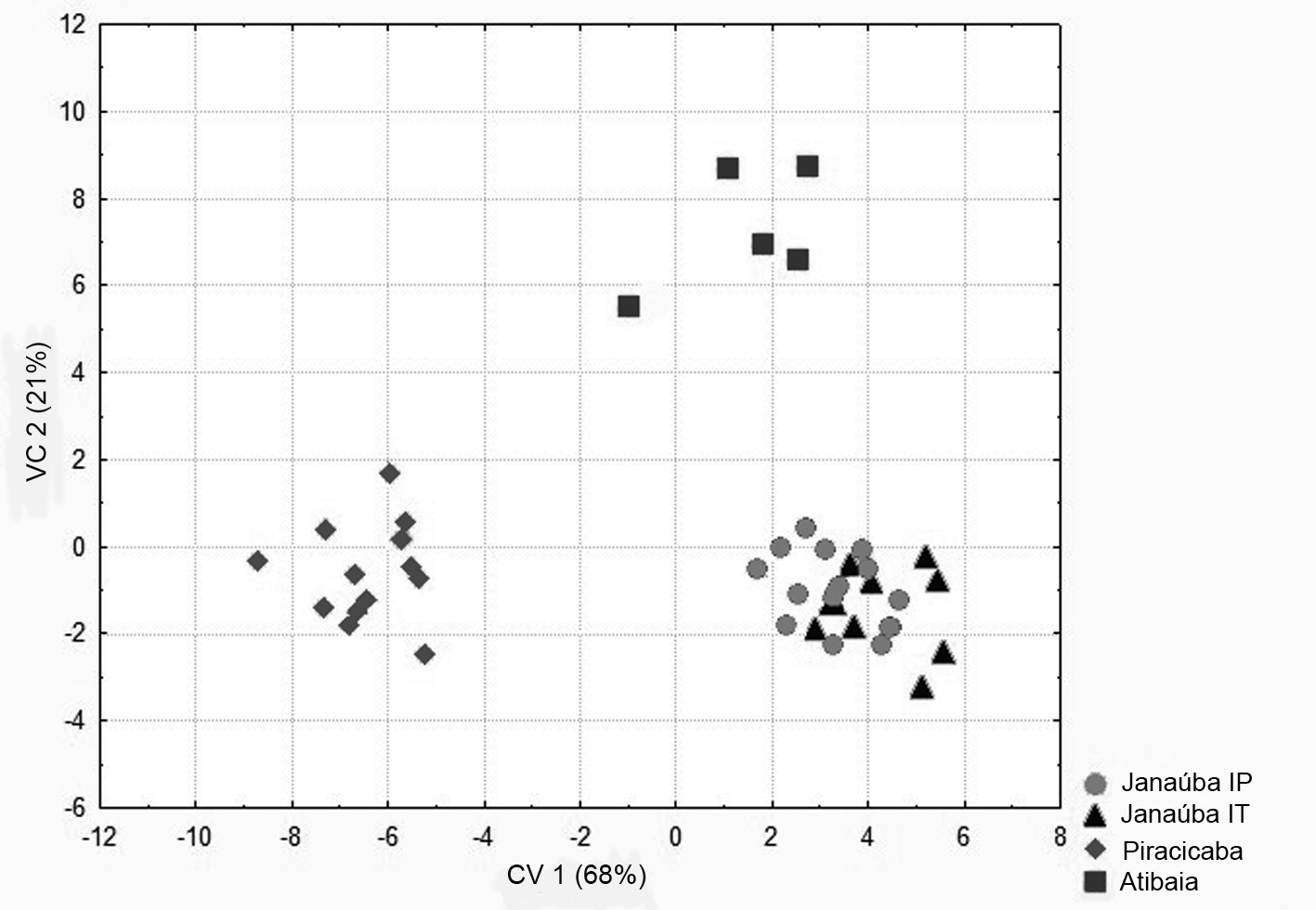
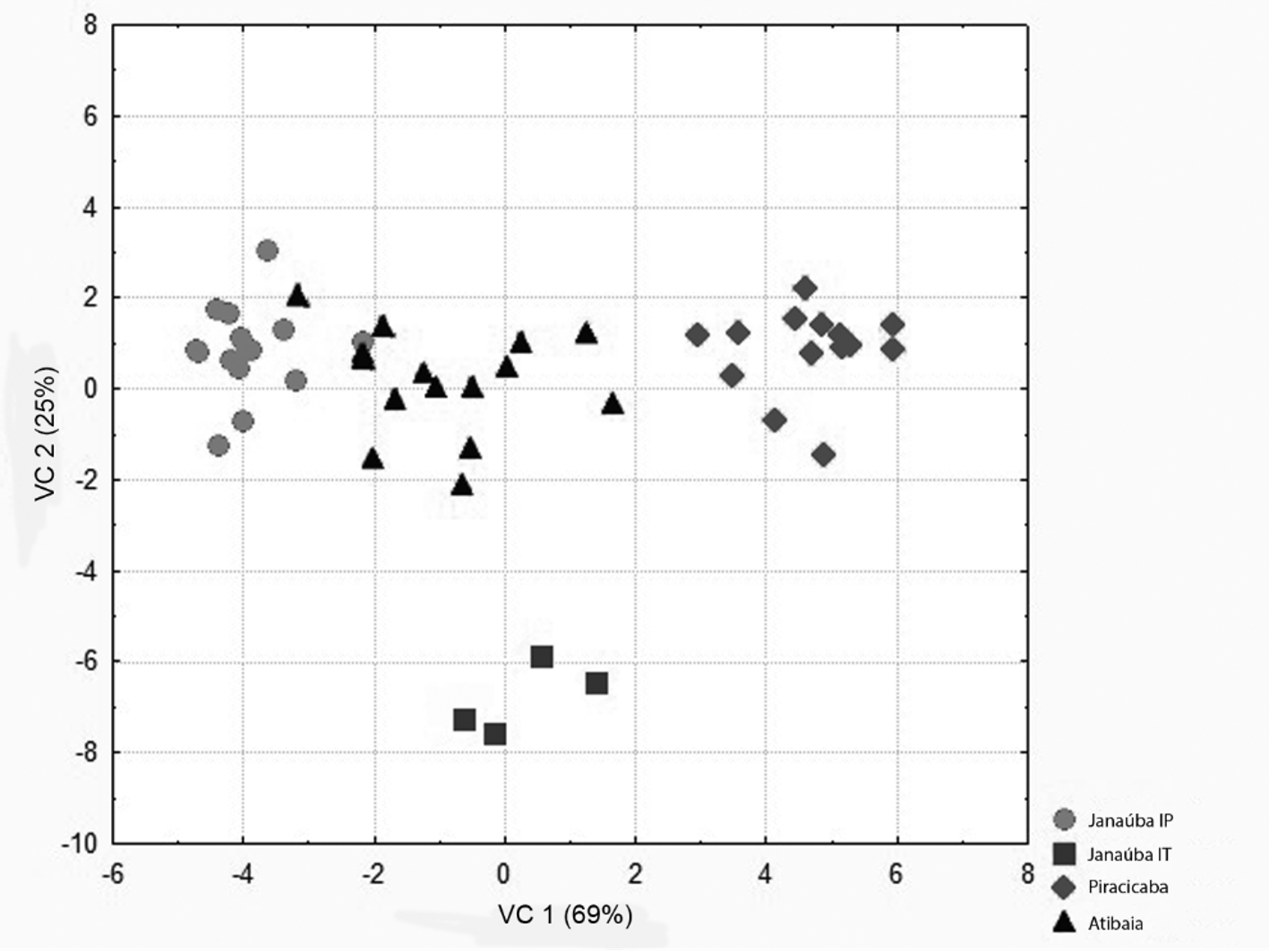
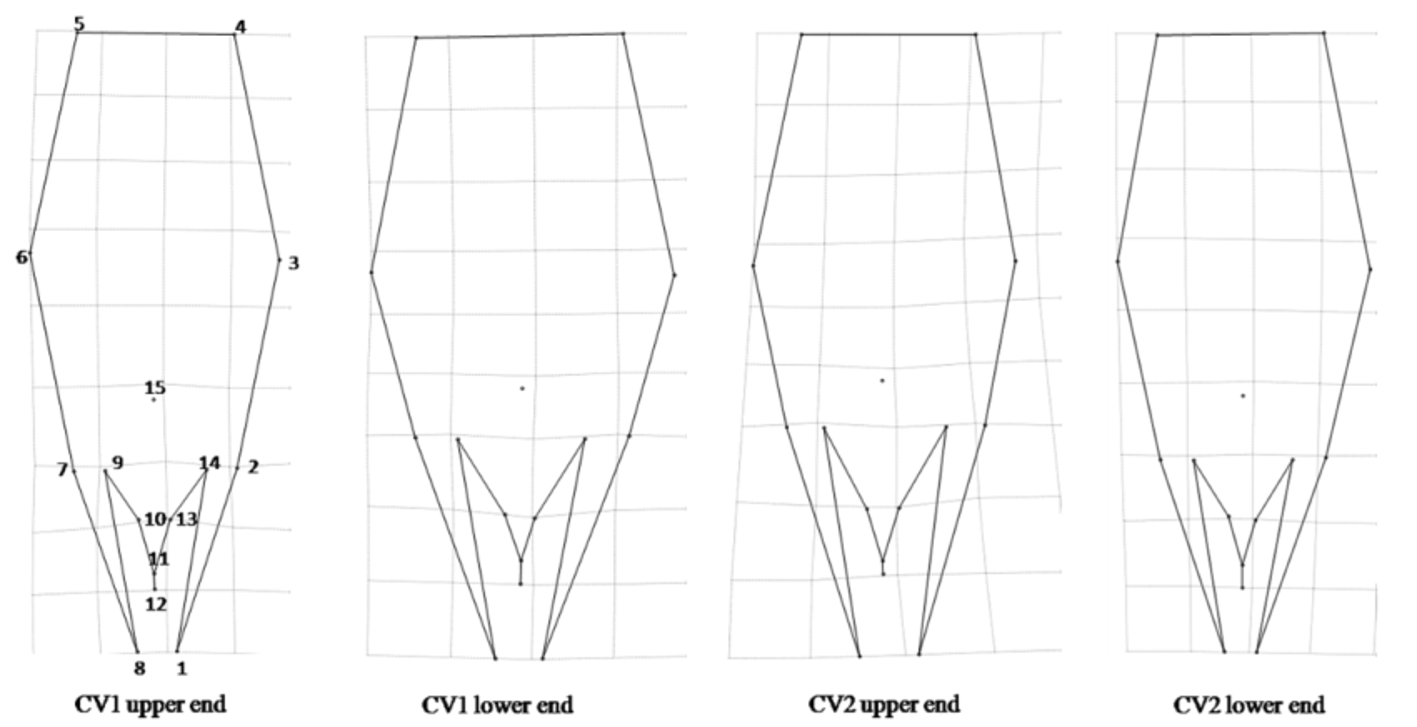
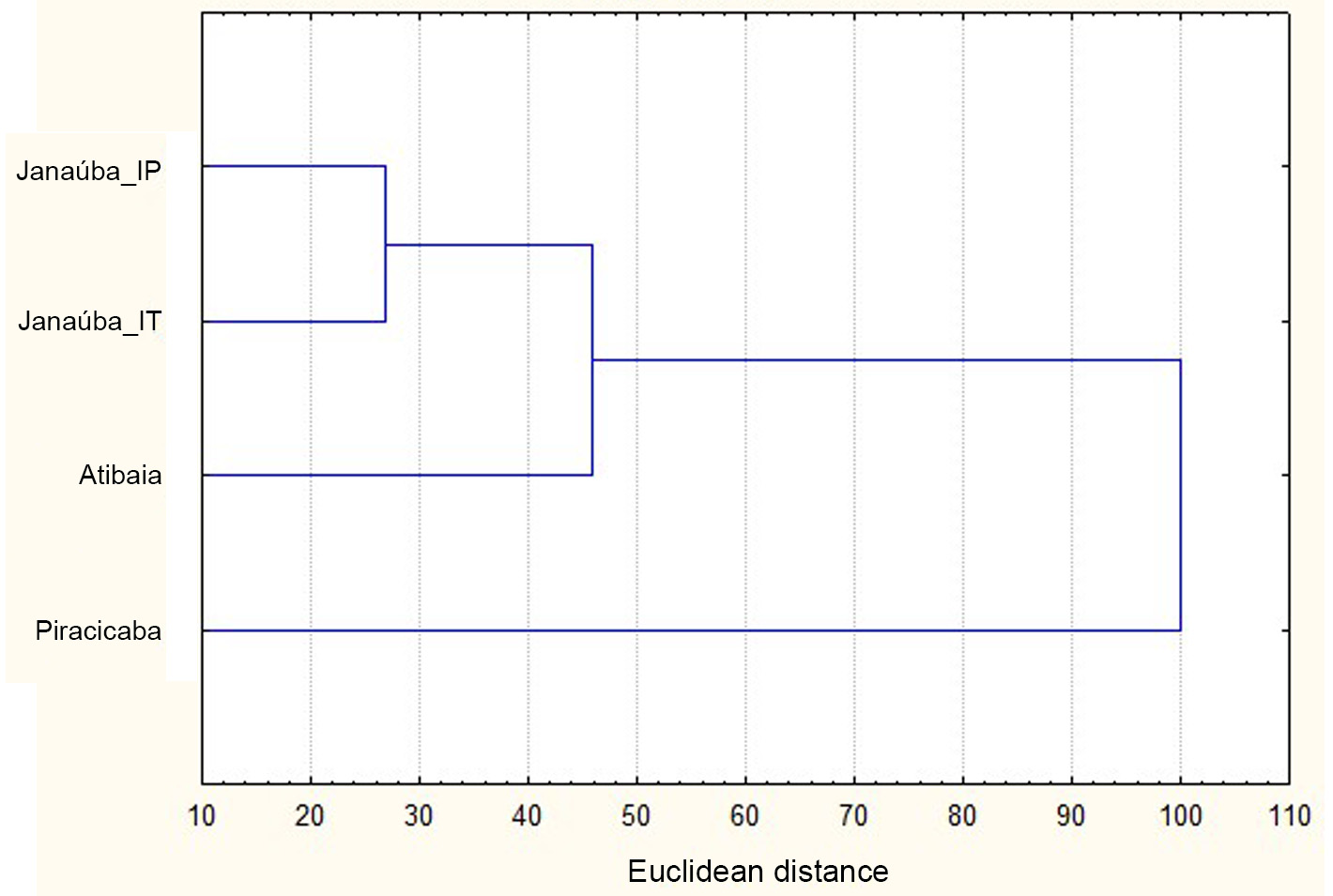
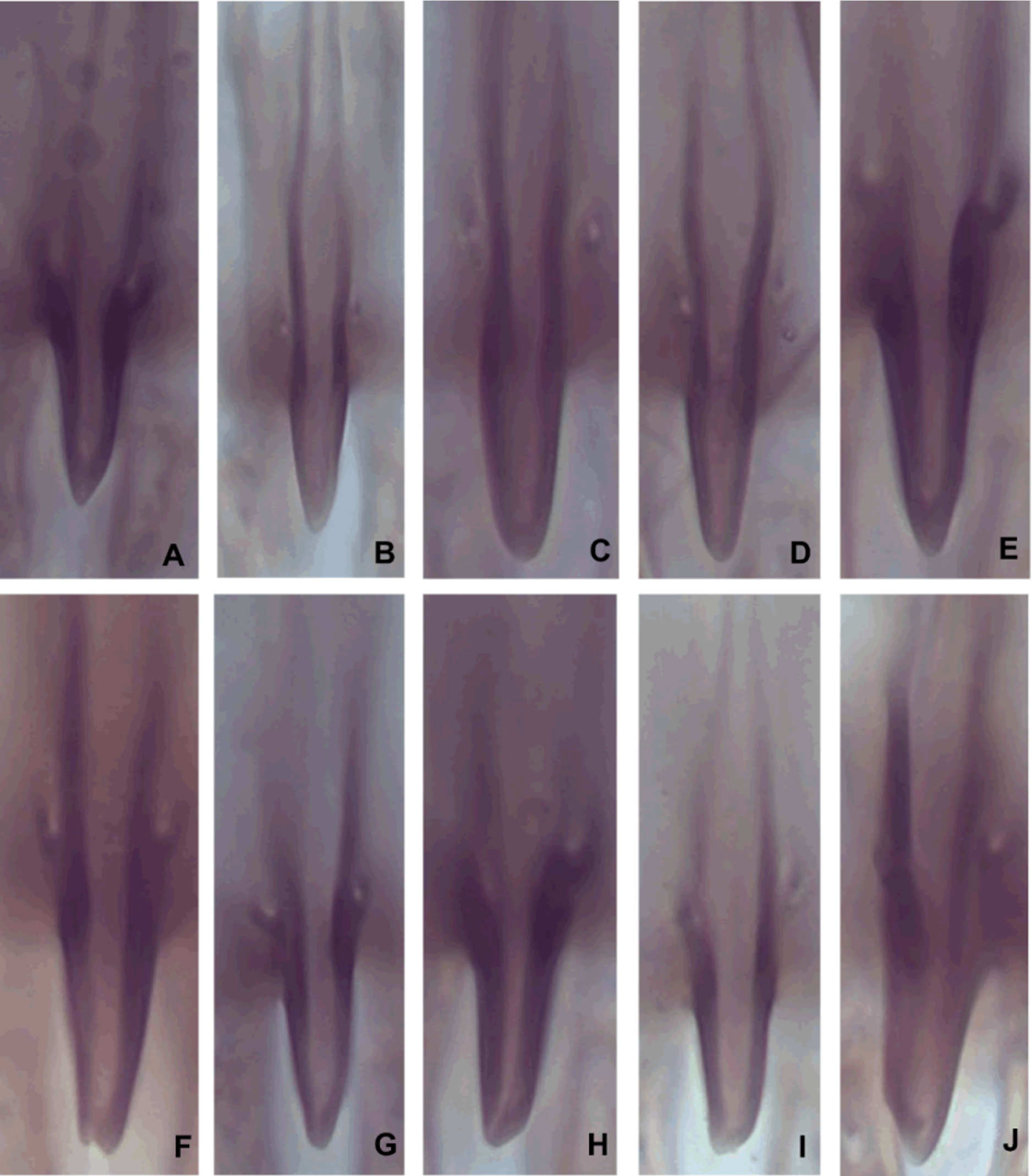
TABLE 2. Mean number of offspring (male and female) obtained from crosses of Trichogramma marandobai with IT and IP processes, reared on eggs of Heliothis virescens at ± 25 ° C.
| Crosses | n | males | females | Total | Parasitizedeggs |
|---|---|---|---|---|---|
| Heterogamic | |||||
| ♂ IP x ♀ IT | 25 | 1.00 ± 0.32 | 2.48 ± 0.93 | 3.48 ± 1.20 | 4.76 ± 0.94 |
| ♂ IT x ♀ IP | 28 | 0.61 ± 0.29 | 3.68 ± 1.37 | 4.29 ± 1.62 | 5.11 ± 0.96 |
| Homogamic | |||||
| ♂ IP x ♀ IP | 21 | 0.81 ± 0.38 | 2.10 ± 1.11 | 2.90 ± 1.38 | 3.62 ± 0.91 |
| ♂ TMT x ♀ TMT | 28 | 1.00 ± 0.36 | 3.11 ± 1.19 | 4.11 ± 1.51 | 5.5 4± 1.11 |
| Parthenogenesis | |||||
| ♀ IP virgin | 19 | 1.68 | 0 | 1.68 | 2.58 |
| ♀ IT virgin | 43 | 9 | 0 | 9 | 6.14 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Trichogramma marandobai Brun, Moraes & Soares, 1986
| Vieira, Jaci Mendes, Querino, Ranyse B., Cônsoli, Fernando L. & Zucchi, Roberto A. 2015 |
Trichogramma demoraesi:
| Querino 2011: 38 |
| Zucchi 2010: 231 |
| Querino 2005: 51 |
| Teles 2005: 515 |
| Zucchi 1997: 53 |
| Zucchi 1985: 43 |