
Rioneta Johnson, 1975
|
publication ID |
https://doi.org/10.5281/zenodo.181365 |
|
DOI |
https://doi.org/10.5281/zenodo.6231924 |
|
persistent identifier |
https://treatment.plazi.org/id/03C2B611-FFB7-A76E-FF3D-FE7DFC1BF9B6 |
|
treatment provided by |
Plazi |
|
scientific name |
Rioneta Johnson, 1975 |
| status |
|
Rioneta Johnson, 1975 View in CoL
Johnson 1975: 719 (generic description) Hall 2000: 240 (list of Ptiliini genera)
Type species. Rioneta uluguruensis Johnson, 1975 , by original designation.
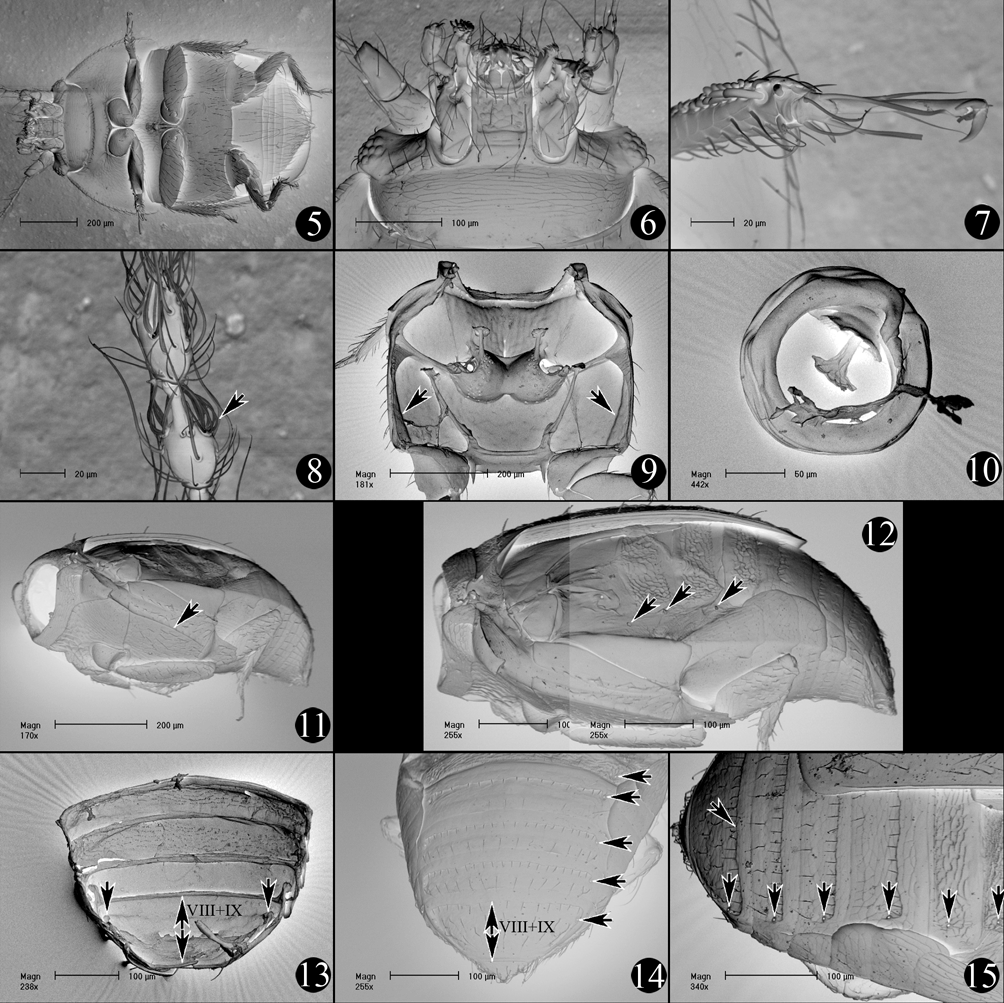
Diagnosis. The genus Rioneta can be distinguished from the majority of other Ptiliidae by the complete absence of hind wings and some correlating modifications of the pterothorax and abdominal tergites such as a markedly transverse scutellum ( Fig. 23 View FIGURES 23 – 33 ), reduced ridges of the metathorax ( Fig. 24 View FIGURES 23 – 33 ), the absence of fingerprint-shaped wing-folding patches on abdominal tergites II–V ( Fig. 15 View FIGURES 5 – 15 ) and the highly reduced or absent palisade fringe on the posterior edge of abdominal tergite VII ( Fig. 15 View FIGURES 5 – 15 ). From the few other wingless genera of Ptillidae (members of the subfamily Cephaloplectinae distributed in the Neotropical and Australian Regions and the monotypic genus Malkinella Dybas, 1960 restricted to the Cape Province of South Africa) the genus Rioneta is easily distinguished by the presence of complete lateral longitudinal grooves on the metaventrite ( Fig. 11 View FIGURES 5 – 15 ) and by the unique ring-shaped spermatheca with the sperm pump located inside the ring ( Figs 1, 4 View FIGURES 1 – 4 , 10 View FIGURES 5 – 15 , 29, 30 View FIGURES 23 – 33 , 34, 35 View FIGURES 34 – 36. 34, 35 ).
Description. General appearance. Body large (1.0 mm and larger); dorsal body surface dull, not shiny; dorsal body pubescence with many short recumbent setae; elytral setae not forming longitudinal rows; body neither markedly flattened nor thickened in vertical dimension; body greyish; color of antennae similar to that of body; wings absent; wing polymorphism absent or unknown. Head. Posterior of eyes without transverse groove or foveae dorsally; frons without protuberances; head capsule with transverse ridge originating from maxillary base and extended laterad, straight, oblique, single, not reaching eyes; temples behind eyes without transverse carina; genal ridge to accommodate antennae in repose absent; transverse row of pore-like structures on neck absent ventrally; medial projection of tentorial bridge absent or inconspicuous; tentorial bridge moderately bent at middle ( Fig. 4 View FIGURES 1 – 4 ); posterior edge of eyes not abruptly cut in frontal plane, without concavity to accommodate pronotum; eyes present, markedly reduced in size, not markedly protruding; interfacetal setae present in posterior part of eyes; antennae of medium length, with 11 antennomeres; with their apices reaching hind pronotal edge; antennomere 3 not widened proximally; antennomeres 4–7 of medium length, about 3–4 times as long as wide; antennal club loosely delimited, composed of 3 antennomeres; apical antennomere not dumbbell-shaped; basal ring present on antennomeres 8–11; apical part of each of two penultimate antennomeres clearly narrowed and elongated into tube; modified thickened setae on two apical antennomeres present, organized in 2 and 3 groups of 5–7 setae ( Fig. 8 View FIGURES 5 – 15 ). Pronotum. Dorsal pronotal grooves absent; short longitudinal ridges on pronotum absent; prothoracic hind angles rounded, not pointed posteriorly, not or slightly expanded posteriorly; lateral ridges on pronotum present, fully separating dorsal and lateral surfaces of pronotum; lateral margins of pronotum not serrate, without sinuation in basal half; anterior edge of pronotum markedly narrower than posterior; posterior edge of pronotum straight at middle; ventral prosternal process absent. Pterothorax. Elytra short and truncate, exposing at least tergites VII to X; medial and posterior edges of elytra without microserration; hind wings absent; groove of alacrista with microserration; alacrista without single seta-like spur in posterior part (= metascutellar spur) absent; mesocoxal acetabula narrowly separated by less than 1/4 acetabular diameter; mesoventral collar clearly delimited, extending laterally to shoulders; disc of mesoventrite (=mesosternal elevation) with longitudinal carina along midline; disc of mesoventrite without clearly contrasting dark spot at centre; disc of mesoventrite not spatulate; posterior apex of mesoventral intermesocoxal process extending beyond middle of mesocoxal acetabula; mesoventral foveae absent; meso-metaventral suture straight, directed laterad and anteriorly, not indented, 1.5X times as long as diameter of mesocoxal acetabulum; lateral longitudinal keel on each side of metaventrite (not to be confused with metaventral lines) present and reaching posterior edge of metaventrite ( Fig. 11 View FIGURES 5 – 15 ); metendosternal arms internally moderately divergent; metendosternite not forked; metacoxae moderately separated by less than twice diameter of mesocoxal acetabulum; metaventrite with posteriorly oriented teeth mediad to metacoxae; metaventral lines absent; metathoracic episterna not or vaguely visible in ventral view; hind femoral plates (=metacoxal laminae) large and subequal in surface size to mesocoxal acetabula; suture connecting outer point of metacoxal acetabulum with internal hind angle of metepisternum long, about as long as metacoxal anterior edge. Abdomen. Laterotergites present on abdominal segments III–VII, prominent, covering margins of respective tergites; palisade fringe along posterior margin of tergum VII markedly reduced in size or absent; fingerprint-shaped groups of wing-folding microridges on tergites II–V absent; pair of abdominal glands present ( Fig. 26 View FIGURES 23 – 33 ); tergite IX without granular microsculpture, without transverse row of pores basally; tergites VIII and IX fully fused to form a single plate ( Figs 13 View FIGURES 5 – 15 , 27 View FIGURES 23 – 33 ); tergite X discrete; pygidial hind margin with single obtuse apical projection, without fan-shaped group of posteriorly oriented setae; spiracular opening present on tergite VIII; abdominal sternite VIII without cavities; abdominal sternite III without longitudinal keels (=femoral lines); length of abdominal sternites VIII 1 –1.5X that of any of sternites III–VII; male abdominal sternite IX with anteriorly directed handle-like projection ( Fig. 33 View FIGURES 23 – 33 ); aedeagus oriented asymmetrically in abdomen, more than 2X as long as wide ( Fig. 31 View FIGURES 23 – 33 ); parameres absent; spermatheca ring-shaped ( Figs 10 View FIGURES 5 – 15 , 29, 30 View FIGURES 23 – 33 ).
Monophyly. As currently defined, the genus Rioneta is almost certainly monophyletic. Both of its species are similar morphologically with the body proportions being the only feature to differentiate them ( Figs 1–4 View FIGURES 1 – 4 ). Both species share a number of morphological traits, which are either rare, or unique in Ptiliidae , which strongly suggests monophyly of the genus. These characters are: (01.) aptery, with complex of associated morphological modifications (see below), (02.) lateral longitudinal keel on each side of metaventrite ( Fig. 11 View FIGURES 5 – 15 ), (03.) fully fused abdominal tergites VIII and IX ( Figs 13, 14 View FIGURES 5 – 15 , 27 View FIGURES 23 – 33 ), (04.) shape of markedly elongate abdominal glands with long duct ( Fig. 26 View FIGURES 23 – 33 ), and (05.) unique shape of spermatheca forming a closed ring with sperm pump located in same surface inside the ring ( Figs 29, 30 View FIGURES 23 – 33 ).
Biology. Both species of Rioneta were collected from forest leaf litter in wet and apparently primary tropical rainforest at altitudes between 2,000 and 2,500 meters. Guts of at least some Euparal-mounted specimens contained fungal spores ( Fig. 1 View FIGURES 1 – 4 ).
Distribution. As currently known, the genus Rioneta is strictly endemic to the Uluguru Mountains in central Tanzania, a part of the East Arc mountain chain. Both known collecting records ( Johnson, 1975 and the present one) are from the southern part of the Ulugurus on the western slope of Lukwangule Plateau ( S07°07’33” E037°36’16”) just above the village of Tchenzema.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.