
Asphinctopone pilosa Hawkes
|
publication ID |
https://doi.org/ 10.5281/zenodo.195422 |
|
DOI |
https://doi.org/10.5281/zenodo.6494201 |
|
persistent identifier |
https://treatment.plazi.org/id/03BB87B9-FF9B-0E2E-FF14-135C0942FE47 |
|
treatment provided by |
Plazi |
|
scientific name |
Asphinctopone pilosa Hawkes |
| status |
sp. nov. |
Asphinctopone pilosa Hawkes sp. n.
( Fig. 1 View FIGURE 1 , A–E)
Holotype worker. TL 4.48, HL 1.07, HW 0.86, CI 81, SL 0.80, SI 92, OD 0.06, OI 7, PW 0.66, WL 1.38 (all measurements in mm).
Medium sized (total length c. 4.5 mm) ponerine ant largely conforming to the diagnosis of the genus as presented by Bolton & Fisher (2008), but with differences in the development of the promesonotal and metanotal grooves, antennal structure and median clypeal lobe as indicated below.
Mandibles smooth and shining, without any sculpture other than coarse scattered hair-pits and with five teeth plus a small denticle between the basal and the second tooth. No tooth-like process on the inner basal margin of the mandible. Eyes very small, comprising a single central ommatidium surrounded by a ring of 6 less distinct ommatidia, positioned distinctly in front of the mid-length of the sides of the head. Ocular diameter less than half the maximum width of the antennal scape. Entire dorsum, sides and ventral surfaces of head with quite coarse punctures (up to 0.03 mm diameter) with weak irregular sculpture between the punctures on the dorsal and dorso-lateral surfaces. Median portion of the vertex, as well as the sides of the head below the eyes, with larger and more widely spaced punctures. Pubescence on head dense, decumbent and arising both from within the numerous large punctures and from minute hair-pits between these, more sparse on the sides of the head below the eye and on the underside, giving these areas a smoother and more polished appearance. Frontal lobes somewhat more densely and more finely punctulate than remainder of the head; frontal carinae and antennal scrobes absent. In full-face view the sides and the posterior margin of the head very shallowly convex. Antennal scapes when laid back just reaching the occipital margin. Funicular segments 2–10 broader than long, segments 2–7 very distinctly so. Funicular segments 8–11 broader than the preceding 6, becoming increasingly longer relative to their width and forming an indistinctly 4-segmented club. The terminal funicular segment much longer than broad and distinctly exceeding the combined length of the preceding 5 segments. Clypeus with a projecting median lobe bounded by a rounded obtuse angle on each side, the anterior margin of this lobe flat to very slightly concave laterally and with a broadly rounded median projection. Width of the anterior margin of the projecting clypeal lobe about 0.33 x HW. Several pairs of strong setae and a number of weaker setae present on the clypeus.
Mesosoma with a pattern of sculpturing very similar to that of the head; dorsum and sides of pronotum, mesonotum and propodeum entirely coarsely punctulate and covered with a dense pelt of fine suberect (pronotum and mesonotum) to erect (propodeum) curved pubescence, the katepisternum and the disc of the pronotum with more sparse punctures and pilosity. Ventral surfaces of the mesosoma with dense fine puncturation and reduced pubescence. The punctulate sculpture on the dorsal and lateral mesosoma is overlaid by irregular effaced fine rugulose sculpture everywhere except on the disc of the pronotum and small areas of the lower propleuron and katepisternum. Pronotal humeri broadly rounded; the pronotum with a subglobose appearance in dorsal view. Promesonotal suture moderately impressed and with cross-ribs on the anterior mesonotum, the metanotal groove only slightly impressed and with fewer more widely spaced but much longer cross-ribs. Mesonotum twice as wide as long. Propodeal dorsum narrow, about half the width of the mesonotum and extending horizontally from the level of the metanotal groove to the short upper portion of the declivity, which slopes downward at about 45o to the junction with the very steep main face of the declivity. The propodeal outline thus presents three distinct planes in profile view. Propodeal dorsum about 1.5 x as long as the mesonotum. Declivity of the propodeum the only portion of the dorsal mesosoma lacking sculpture and with greatly reduced pilosity; smooth and weakly shining.
Petiole node high and squamiform, unsculptured apart from minute hair-pits, smooth and weakly shining and with pubescence similar to but less dense than that on the mesosoma. In profile view the anterior and posterior faces of the petiole node almost parallel, only slightly convergent dorsally; in posterior view the dorsal margin forming a bluntly triangular peak. In dorsal view the anterior face shallowly convex, the posterior even more shallowly concave. Subpetiolar process with the complex shape characteristic of the genus.
Gaster unsculptured apart from minute hair-pits (c. 0.005 mm diameter), pubescence denser but shorter (0.04 mm) on tergites 2–4 than on the first gastral tergite (0.07 mm), much longer and more variable on the sternites. No setae present on gastral tergites 1–4, but a few pairs on each sternite and abundant setae present on the pygidium and hypopygium. Sting stout and slightly upcurved in lateral view, somewhat laterally flattened.
A single (pectinate) spur is present on each tibia, those on the metatibiae much longer (0.34 mm) than those on the mesotibiae (0.17 mm), with those on the protibiae intermediate in length (0.29 mm). All segments of the appendages, including the antennae, with fine appressed pubescence but no standing hairs.
Colour: head and mesosoma dark reddish-brown throughout, the gaster, appendages and mandibles a lighter reddish-brown, with the exception of a small yellowish-brown patch on each mandible.
Holotype worker. Tanzania, Tanga Region, Kilindi Forest Reserve, 1015 m, S 5.57934 E 37.57971, 28.viii.2005, CEPF-TZ-3.4.F34, SAM-HYM-C 020683, Primary forest, hand collected (P. H a w k e s, J. Makwati, R. Mtana) ( SAMC).
Comments. Asphinctopone pilosa is distinctly different from its congeners in many respects; the most obvious differences are 1) its larger size, 2) the stronger and much more extensive sculpturation and denser pubescence, 3) the differently shaped mesonotum and propodeum, 4) the far less strongly impressed promesonotal suture and metanotal groove, 5) the terminal four, rather than three, antennal segments forming a weak club, 6) the more strongly squamiform petiole, which is also more arcuate in dorsal view, 7) the lessdeveloped clypeal structure (in sylvestrii and differens the median clypeal lobe is bounded by distinct sharp angles, is distinctly though shallowly concave on either side of the more acutely rounded median projection and is relatively broader at about 0.40 x HW), 8) the lack of a tooth-like process on the inner basal margin of the mandible and 9) its darker colour.
Etymology: The specific epithet pilosa refers to the pubescence which is much denser on the head and mesosoma than in all previously described species of the genus.
Habitat. The Kilindi Forest Reserve (5395 ha) lies about 140 km inland and comprises a small area of forest on an isolated double-peaked mountain surrounded by miombo woodland; the survey site was located at about 1000 m altitude on a mainly flat area on the saddle between the two peaks. The forest canopy here was estimated to average about 25 m high, with some emergent trees of 35–40 m; closure of the canopy was estimated at approximately 70–80%. Previous disturbance and tree felling by informal gold miners was evident and numerous young trees and climbers were observed in partially cleared areas. Soils along the survey transect were mainly of sandy loam texture (with some patches of sandy clay loam), a pH of 8.0–8.5 and with a thin (1–2 cm depth) leaf litter layer ranging from 25% to 100% cover. The survey was carried out toward the end of the cool dry season in late August 2005; soil temperatures averaged 16o C at a depth of 10– 15 cm, with soil moisture at the same depth averaging 11.9%. The Asphinctopone pilosa specimen was, however, collected in a microhabitat with higher moisture levels and a much thicker litter layer than average for the site. Humidity during the three-day site visit ranged from 63.4% to 99.3% (the latter during an unseasonal rainstorm) and averaged 81.4%, while the temperature varied from 12.0–22.5o C.
Climate. Detailed climatic data are not readily available for all of the localities from which other Asphinctopone specimens have been collected, but inspection of Worldclim (version 1.3) 2.5 arc-minute extrapolated climate data ( Hijmans et al. 2005) downloaded via http://www.diva-gis.org/climate provides some indication of conditions within the known distribution (see Figure 2 View FIGURE 2 ) of the genus. Most of the localities exhibit low seasonal variation in temperature and precipitation. The A. pilosa locality was an outlier in respect of some of the bioclimatic variables (it has the highest temperature seasonality, lowest mean annual temperature and lowest mean temperature of the driest quarter), but in most variables was similar to previously recorded Asphinctopone localities.
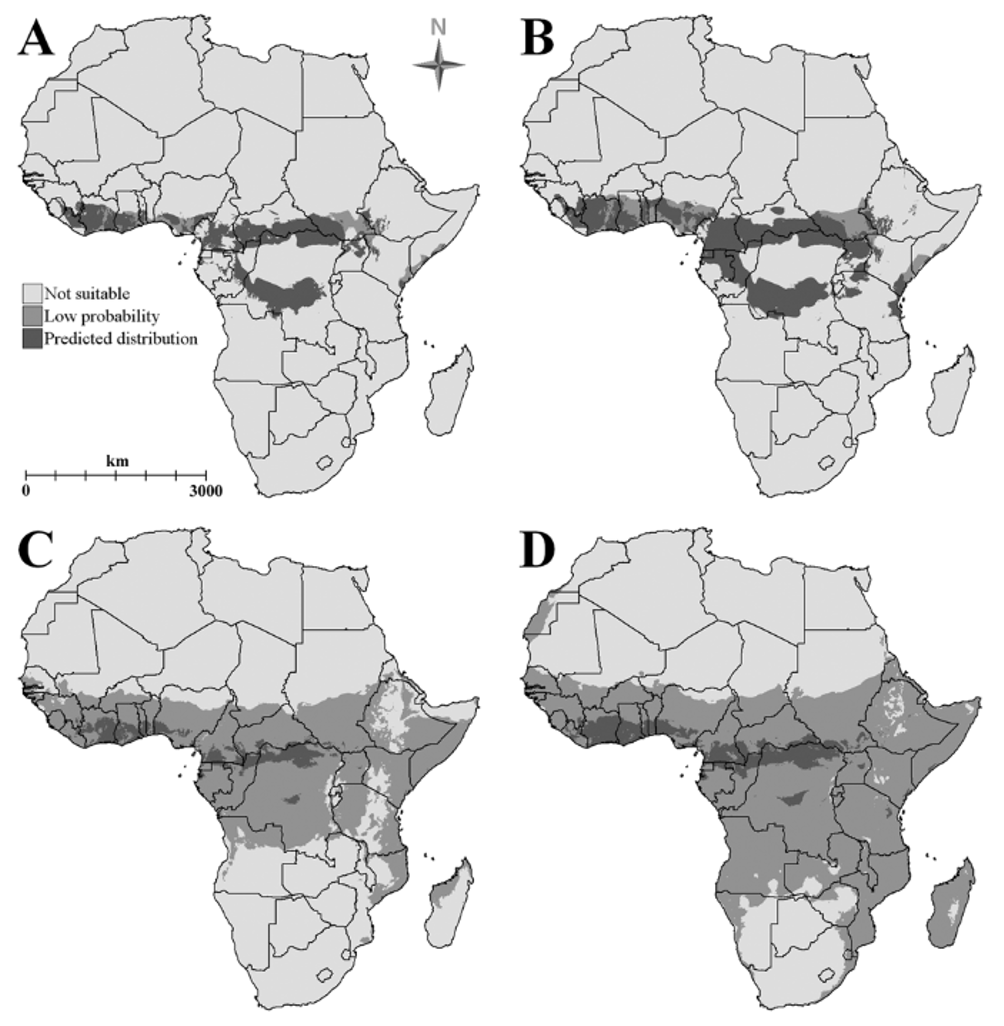
Modelling of habitat suitability and predicted distribution. Of the nine bioclimatic variables selected for inclusion in the model, two showed a close to normal and one a truncated normal distribution, while six were strongly skewed. For three of the variables with skewed distributions (min. temperature of coldest month, mean temperature of coldest quarter and mean temperature of driest quarter) estimates from visual inspection indicated that less than 5% of the entire African continent has values higher than the modal value of the Asphinctopone localities. This suggests that intolerance to low temperatures may be a significant factor influencing distribution. Test predictions in BIOCLIM using all of the 19 bioclimatic variables or the selected subset of nine without the tail box adjustment gave implausible results with most (14–16 of 19) recorded A. silvestrii localities placed at or near (<10 km) an edge of the predicted distribution; using the adjustment and the selected variable subset gave far more probable results, with the majority (13) of these records falling well within the predicted distribution.
In addition to the seven countries ( Cameroon, Central African Republic, Gabon, Ghana, Guinea, Ivory Coast and Nigeria) from which A. silvestrii and A. differens have been recorded to date, BIOCLIM modelling predicted a reasonable probability (see Figure 3 View FIGURE 3 A) that one or both of these species could occur in a further 12 African countries ( Angola, Benin, Democratic Republic of the Congo, Ethiopia, Equatorial Guinea, Kenya, Liberia, Republic of the Congo, Sierra Leone, Sudan, Toga and Uganda). However, in some cases the distribution within these countries is expected to be very limited (e.g. to the far north-east in Angola, the southern Sudan and western Ethiopia). Inclusion of the A. pilosa locality in the model resulted in the predicted suitable area in many of these countries increasing substantially (see Figure 3 View FIGURE 3 B) and also in the inclusion of parts of north-western Tanzania. While this may not be a realistic expansion of the predicted ranges of A. silvestrii and A. differens , it could indicate the possibility of additional Asphinctopone species in areas beyond the ranges of the known species.
It is recognised that BIOCLIM often under-predicts species distributions ( Ward 2007), but the use here of a selected subset of bioclimatic variables and adjusting tail checkboxes resulted in a predicted distribution greater than the core area predicted by DOMAIN ( Figure 3 View FIGURE 3 C and 3D). The large non-core area mapped by the latter model illustrates a low predicted probability of occurrence, so the apparently very large predicted distributions in Figure 3 View FIGURE 3 C and 3D should perhaps be viewed with some scepticism, especially as the latter includes some very improbable areas such as a large portion of the Namib desert.


| SAMC |
Iziko Museums of Cape Town |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Ponerinae |
|
Genus |