
Chiroderma trinitatum Goodwin, 1958
|
publication ID |
https://doi.org/10.11646/zootaxa.4846.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:6F6EBF63-5598-416C-8694-14C4A8687693 |
|
DOI |
https://doi.org/10.5281/zenodo.4332645 |
|
persistent identifier |
https://treatment.plazi.org/id/03BAA52E-F869-FFFB-7090-FB667EC359FD |
|
treatment provided by |
Felipe |
|
scientific name |
Chiroderma trinitatum Goodwin, 1958 |
| status |
|
Chiroderma trinitatum Goodwin, 1958 View in CoL
Synonyms:
Chiroderma trinitatus Goodwin, 1958:1 View in CoL ; type locality “ Cumaca , Trinidad, British West Indies. ”
Chiroderma trinitatum: Handley, 1960:466 View in CoL ; correct gender concordance.
Chiroderma trinitatum trinitatum: Barriga-Bonilla, 1965: 247 View in CoL ; name combination.
Chiroderma trinitratum Linares and Moreno-Mosquera, 2010: 275 ; incorrect subsequent spelling of Chiroderma trinitatum Goodwin, 1958 View in CoL .
Type Material. The holotype, designated in the original publication, is specimen AMNH 175325, a female pregnant when collected in a well-lit cave, by L. Venus and B. Smith on March 22, 1956, in Cumaca, Trinidad and Tobago. The skin is preserved in fluid and nearly all hair has fallen out. There is a transversal cut on the abdomen. The skull has been removed and is in good condition, with all the teeth and cranial bones preserved. The fetus, removed from the type, has a distinct median dorsal stripe and a crown-rump length of approximately 20 mm.
Distribution and Habitat. Specimens are known form Guyana; Suriname; French Guiana; Trinidad; northern, central and western Brazil; northern and eastern Bolivia; eastern Perú; eastern Ecuador; southern and eastern Colombia; and southern Venezuela ( Fig. 23 View FIGURE 23 ).
Records are from humid tropical forests, in the Amazon basin, Orinoco basin, and the Guianas. Some records are from ecotonal areas between humid forests and drier, more seasonal formations, such as in Serra do Roncador (central Brazil), and in San Ramón (northeastern Bolivia). C. trinitatum have been collected from approximately 20 m above sea level (e.g. Belém, Brazil) to elevations near 1,050 m, in the Andean foothills (e.g. Santa Bibiana, Perú), corroborating the altitudinal amplitude given in the literature (e.g. Handley 1976; Ascorra et al. 1996; Solari et al. 2006, 2019).
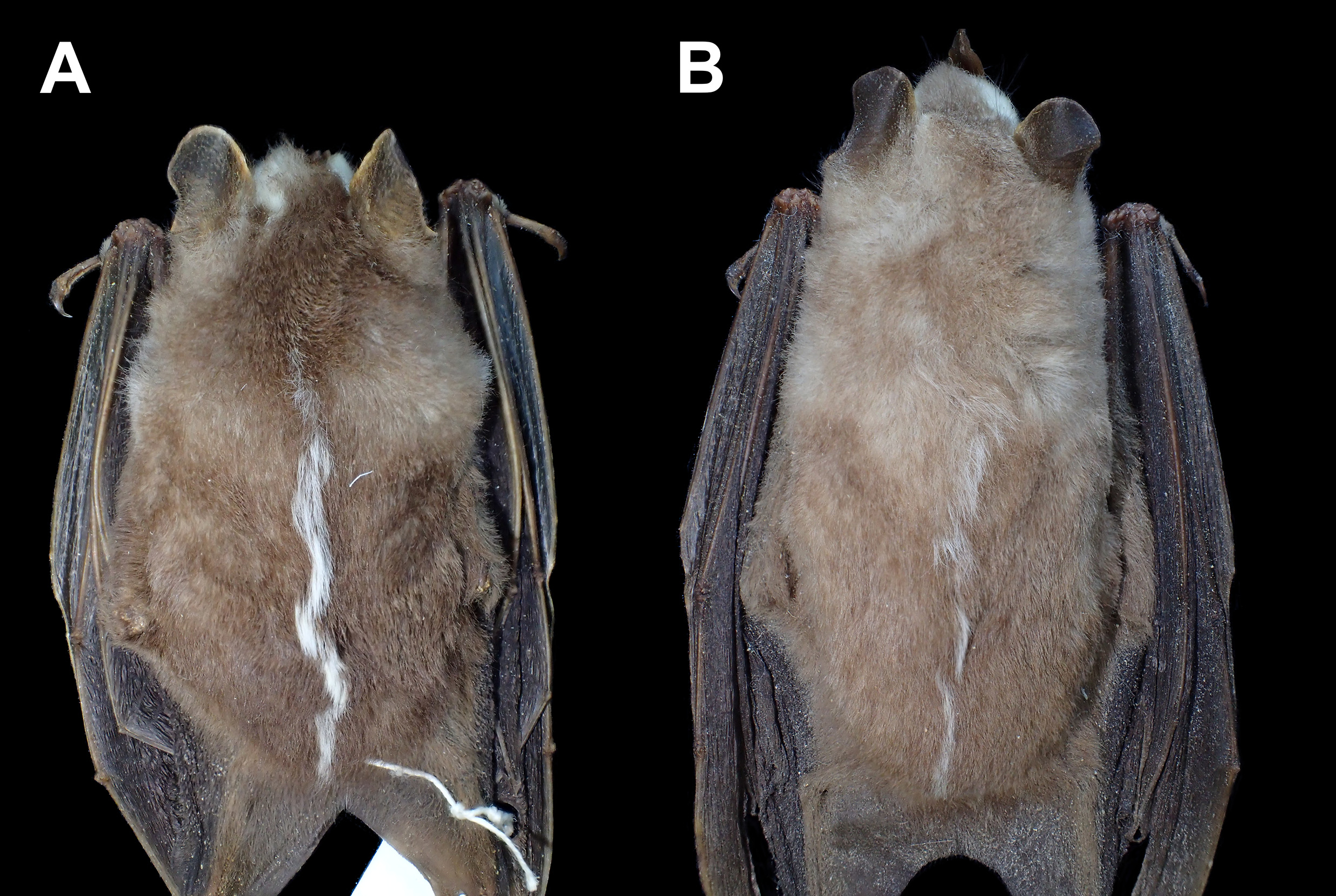
Description and Comparisons. Dorsal pelage may be pale brown, dark brown, or grayish brown ( Fig. 24 View FIGURE 24 ). A completely white C. trinitatum was recorded in the Peruvian Amazon by Tello et al. (2014). Dorsal hairs are tricolored, the base about ¼ of the hair length and dark brown, middle portion about ½ of the hair length and pale buff or pale gray, and tips about ¼ of the hair length and pale brown, dark brown, or grayish brown. Facial stripes are always present with the interocular pair wider (> 2 mm) than the genal stripes. A median dorsal stripe was present in 111 of the 113 specimens examined (98%). When present, the stripe was barely visible in 13 of 95 specimens (13%). The stripe usually begins in the interscapular region and reaches the posterior rump of the body. In some specimens, the dorsal stripe began more anteriorly in the region immediately behind the nape. The ears have yellowish margins and base, with the remainder brownish. The spear of the noseleaf has a simple tip, is brown in color, with paler lateral borders of the horseshoe.
The skull is similar to that of C. gorgasi and it is smaller than every other species of Chiroderma ( Tables 7 and 8). The braincase is globose, standing above the frontonasal region in lateral view. A sagittal crest was absent in 37 of 126 specimens (29.4%), weakly developed in 86 (68.2%), and well developed in 3 (2.3%). The nasal notch is relatively short, not reaching the interorbital region ( Fig. 25 View FIGURE 25 ). C. trinitatum has relatively small orbits; the anterior border approximating the mesial margin of M1 ( Fig. 26 View FIGURE 26 ). Post orbital processes are present, but rhomboid instead of pointed as in the other species.
A posterior palatine process was absent in 110 (90.9%) of the 121 specimens examined. A small process was present in 10 specimens, and one (AMNH 264076) had an anomalous notch in place of a projection on the margin of the palate. Paraoccipital processes are absent. When cranium and mandible are in occlusion, there is a lateral gap bordered by the C, P3, P4, p2 and p4 ( Fig. 9 View FIGURE 9 ).
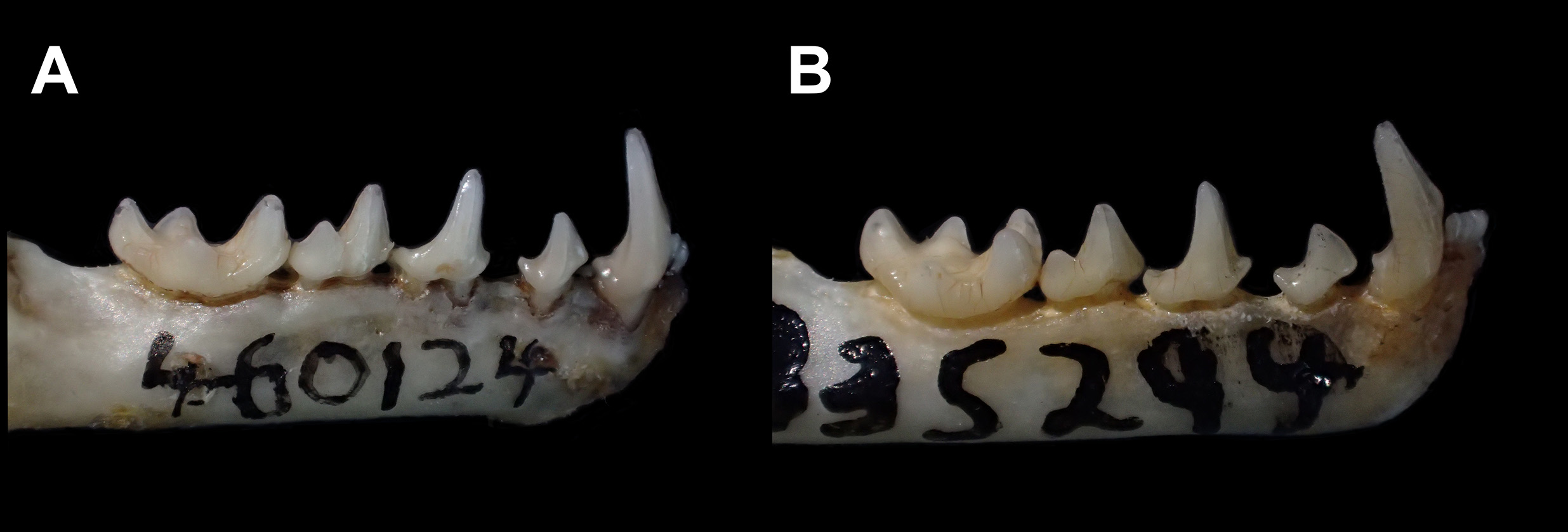
The I1s have converging tips in most of the specimens (112 out of 122) we examined. The I1s may be in contact at the base, along approximately ⅔ of their length, or only at their tips, the most frequent state. In some C. trinitatum , the I1s are separated throughout their length. The P3 is wider (buccolingually) than long (mesiodistally) and does not touch P4.
The crown of the lower canine is relatively short, clearly below the level of the tip of the coronoid, in lateral view ( Fig. 26 View FIGURE 26 ). The p2 is large, approximately ¾ of the height of p4, and higher than long. The p2 may be close to or in contact with the canine, or it may lie approximately equidistant between c and p4 ( Fig. 26 View FIGURE 26 ).
Chiroderma trinitatum is easily differentiated from C. doriae and C. improvisum by its much smaller size ( Tables 7 and 8). The intermediate-sized species, e.g. C. salvini , C. scopaeum , and C. villosum , are distinguishable from C. trinitatum by having their longer nasal notch that reaches the interorbital region, pointed post-orbital processes and the short p2, which is approximately ¼ the crown height of p4.
The species most similar to C. trinitatum is C. gorgasi . C. trinitatum has relatively shorter lower canines, the tips of which are clearly below the level of the top of the coronoid process in lateral view. The lower canines have higher crowns in C. gorgasi , being on the same level or higher than the tip of the coronoid process ( Fig. 26 View FIGURE 26 ). The base of lower canines of C. trinitatum is also longer, but shorter in C. gorgasi . The p2 of C. trinitatum is usually higher than long mesiodistally; whereas, in C. gorgasi the p2 is longer than high ( Fig. 27 View FIGURE 27 ). As recently described by Lim et al. (2020), C. trinitatum tends to have a wider braincase and typically has a third cuspid on p4 that is absent in C. gorgasi .
Geographic Variation and Phylogeography. Phylogenetic analyses of 68 sequences of C. trinitatum resulted in two highly-supported clades ( Fig. 21 View FIGURE 21 ). One of the clades is based on sequences from 26 specimens from the Guianas (French Guiana, Guyana, and Suriname), and the island of Trinidad. The other clade consists of 42 specimens, from the Guianas ( Guyana and Suriname) and the Amazon basin ( Bolivia, Brazil, Ecuador, and Perú) ( Fig. 21 View FIGURE 21 ). However, we could not find any morphological differences between the two clades.
Subspecies. C. trinitatum is monotypic.
Natural History. C. trinitatum is frugivorous and is known to consume fruits and infructescences of at least five species: Cecropia obtusifolia , Piper elongatum , Solanum riparium (Solanaceae) , Ficus sp., and Vismia sp. ( Hypericaceae ) ( Reis & Peracchi 1987; Loayza et al. 2006; Linares & Moreno-Mosquera 2010). C. trinitatum has also been recorded drinking the mineral-rich water at clay licks (“collpas”) in the Peruvian Amazon ( Bravo et al. 2008, 2010; Ghanem et al. 2013; Ghanem & Voigt 2014).
In most studies comparing the relative abundance of species in the canopy and understory, C. trinitatum was more commonly netted in the higher forest strata, at approximately 20 m, suggesting that the species is a canopy frugivore ( Ascorra et al. 1996; Simmons & Voss 1998; Charles-Dominique & Cockle 2001; Kalko & Handley Jr. 2001; Delaval et al. 2005; Rex et al. 2011). Day roosts of C. trinitatum are unknown, and the only information available comes from the type specimen, which was collected on the island of Trinidad in a cave described by Goodwin & Greenhall (1961) as well-lit and co-inhabited by Micronycteris megalotis .
In Venezuela, two species of ectoparasites were documented on C. trinitatum , the mite Periglischurus iheringi (Spinturnicidae) and a species of Streblidae of the genus Paratrichobius (salvini complex) ( Herrin & Tipton 1975; Wenzel 1976). In Colombia, the tick Ornithodoros azteci (Argasidae) was recorded from C. trinitatum by Marinkelle & Grose (1981). The endoparasite Hasstilesia tricolor (Nematoda) was recorded by Nogueira et al. (2014) in C. trinitatum from Acre, Brazil.
Reproductive data suggest seasonal polyestry. Pregnant females are recorded from December through March in the Amazon of Colombia, Perú, and Venezuela, and on the island of Trinidad. Pregnancies are recorded from June through September in Guyana and the Amazon of Brazil, Perú, and Venezuela. Lactating females were documented in February, April, May, and July. Apparently, gestation peaks during the rainy season (December to March), with one birth peak from the middle to the end of the rainy season, and a second during the dry season (June to September) with births occurring close to the beginning of the rainy season.
Specimens Examined (N = 146): Bolivia: Beni, Estancia Yutiole ( AMNH 210810 ), Remansos ( AMNH 209520 , 209521 ); La Paz, Río Coraico Valley ( AMNH 246646 ), Santa Ana de Madidi ( AMNH 261632 , 261641 );
Santa Cruz , 10 km N San Ramón ( AMNH 261674 About AMNH ), Parque Nacional Noel Kempff Mercado , 23 km S Campamento Los Fierros ( AMNH 264077 About AMNH ), Parque Nacional Noel Kempff Mercado , 27.5 km S Campamento Los Fierros ( AMNH 264076 About AMNH ), Parque Nacional Noel Kempff Mercado , 3 km S Campamento Los Fierros ( AMNH 264075 About AMNH ), Parque Nacional Noel Kempff Mercado , El Refugio ( USNM 584492 About USNM ). Brazil: Acre, Parque Nacional da Serra do Divisor ( ALP 7020 , 7088 , 7099 , 7124 , 7134 , 7143 , 7144 , 7195 , 7295 , 7311 ); Amazonas, Comunidade Cachoeirinha ( LMSUP [ ICA033 ]), opposite to Comunidade São Pedro ( LMUSP [ ICA173 ]); Mato Grosso, 264 km N Xavantina ( USNM 393704–393711 About USNM ), Parque Nacional do Juruena , Serra dos Apiacás ( CMUFLA 1284 ), São José do Rio Claro ( MZUSP [ PEV 896–897 View Materials ]); Pará, Fazenda Bocaina ( UFMG [ VCT1437 View Materials ]), Fazenda Fartura ( MZUSP 36012 View Materials , 36013 View Materials ), Floresta Nacional de Carajás ( UFMG [ VCT6342 View Materials ]), rio Xingu , Linha de Transmissão Jurupari ( MZUSP 35033 View Materials ), Sta. A , IAN ( USNM 361723 About USNM , 460127 About USNM ), Várzea , Belém ( USNM 460124 About USNM , 460125 About USNM , 460126 About USNM ); Rondônia, Monte Negro ( MZUSP 35026 View Materials , ZUFMS 1342 ). Colombia: Amazonas, Puerto Nariño ( USNM 483766–483769 About USNM ) ; Vichada, Territorio Faunistico Tuparro ( IAvH-M 2083 ). Guyana: Cuyuni-Mazaruni, Maz 3 ( BMNH 1980.751 , 1980.752 ). Namai Creek ( ROM 108144 About ROM ); Potaro-Siparuni, Iwokrama Reserve ( ROM 109026 About ROM ); Upper Demerara-Berbice, 3.2 km W Kurupukari ( BMNH 1997.39 ), Tropenbos ( ROM 103486 About ROM ). French Guiana: Sinnamary, Paracou ( AMNH 266255 About AMNH , 266256 About AMNH , 267189 About AMNH , 268532 About AMNH , 269118 About AMNH ). Perú: Amazonas, Cordillera del Condor ( USNM 581934 About USNM ); Cusco, Camisea , Armihuari ( MUSM 13619 , 13622 , 13623 , USNM 582837 About USNM ), Camisea , Pagoreni ( MUSM 13624–13626 , USNM 582838 About USNM ), Consuelo ( MUSM 19670 ); Huánuco, Puerto Inca ( MUSM 36692 ); Junín, Santa Bibiana ( MUSM 40606 ); Loreto , Jenaro Herrera ( MUSM 4219 , 5594 ), km 22,7 da rodovia Iquitos-Nauta ( MUSM 29559 ), Peña Negra ( MUSM 29557 ), Quebrada Grande ( MUSM 21134 , 21135 ), Río Lagartococha ( MUSM 21138 ); Madre de Dios, Albergue Maskoitania ( MUSM 19669 ), Pakitza ( MUSM 678 ), Quebrada Aguas Calientes ( MUSM 16651 ), Refugio Juliaca ( MUSM 11662 ); Pasco, Campamento Río Lobo ( MUSM 10229 ), Cerro Chontiya ( MUSM 10226 , 10227 ), Cerro Jonatán ( MUSM 10228 ), San Pablo ( AMNH 230656 ), Yanahuanca ( MUSM 10881 ); Puno, Curva Alegre ( MUSM 26658 ), San Fermín ( MUSM 26659 ); Ucayali, Concesión de Conservación Río La Novia ( MUSM 44185 ); Suriname: Brokopondo, Brownsberg Nature Park ( ROM 114213 About ROM ); Sipaliwini, Kushere Landing ( ROM 120168 ). Trinidad and Tobago: Trinidad, Arima ( AMNH 205373 About AMNH ) , Cumaca ( AMNH 175325 About AMNH [holotype of trinitatum ]) View Materials , Fillete ( AMNH 205375 About AMNH ), Las Cuevas ( BMNH 1971.121 ). Venezuela: Amazonas, Boca Mavaca ( USNM 405159 About USNM ), Capibara ( USNM 415245 About USNM ), Cerro Neblina base camp ( USNM 560764–560766 About USNM ), Guayabal ( USNM 415256 About USNM , 415257 About USNM ), Río Cunucunuma ( MZUSP 27166 View Materials , 27167 View Materials , USNM 405132–405139 About USNM , 405141–405148 About USNM , 405156 About USNM , 405157 About USNM ), Río Manapiare ( USNM 415250–415254 About USNM ); Apure, La Blanquita ( USNM 440348 About USNM ); Barinas, Altamira ( USNM 418121 About USNM ); Bolívar, 85 km SSE El Dorado ( USNM 387192 About USNM ), El Manaco ( USNM 387193–387198 About USNM , 387202 About USNM , 387203 About USNM ); Yaracuy, Minas de Aroa ( USNM 440747 About USNM ) .




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Stenodermatinae |
|
Tribe |
Stenodermatini |
|
SubTribe |
Vampyressina |
|
Genus |
Chiroderma trinitatum Goodwin, 1958
| Garbino, Guilherme S. T., Lim, Burton K. & Tavares, Valéria Da C. 2020 |
Chiroderma trinitratum
| Linares, E. L. & Moreno-Mosquera, E. A. 2010: 275 |
Chiroderma trinitatum trinitatum :
| Barriga-Bonilla, E. 1965: 247 |