
Chiroderma gorgasi Handley, 1960
|
publication ID |
https://doi.org/10.11646/zootaxa.4846.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:6F6EBF63-5598-416C-8694-14C4A8687693 |
|
DOI |
https://doi.org/10.5281/zenodo.4332649 |
|
persistent identifier |
https://treatment.plazi.org/id/03BAA52E-F865-FFFC-7090-FAB97D435AFF |
|
treatment provided by |
Felipe |
|
scientific name |
Chiroderma gorgasi Handley, 1960 |
| status |
|
Chiroderma gorgasi Handley, 1960 View in CoL
Synonyms:
Chiroderma gorgasi Handley, 1960: 464 View in CoL ; type locality “ Tacarcuna Village , 3,200 ft., Río Pucro, Darién, Panama.”
Chiroderma trinitatum gorgasi: Barriga-Bonilla, 1965: 246 View in CoL ; name combination.
Type Material. The type of C. gorgasi , USNM 309903 , consists of a stuffed skin, skull and mandible, collected in Tacarcuna Village , Panamá, on March 6, 1959 by C. O. Handley and B. R. Feinstein (field number COHJR 5436). It is an adult male, captured in a mist net over water .
The skin is in good condition with the facial and dorsal stripes observable in the specimen. The skull and mandible are in good condition and every tooth is present. The I1 have convergent tips.
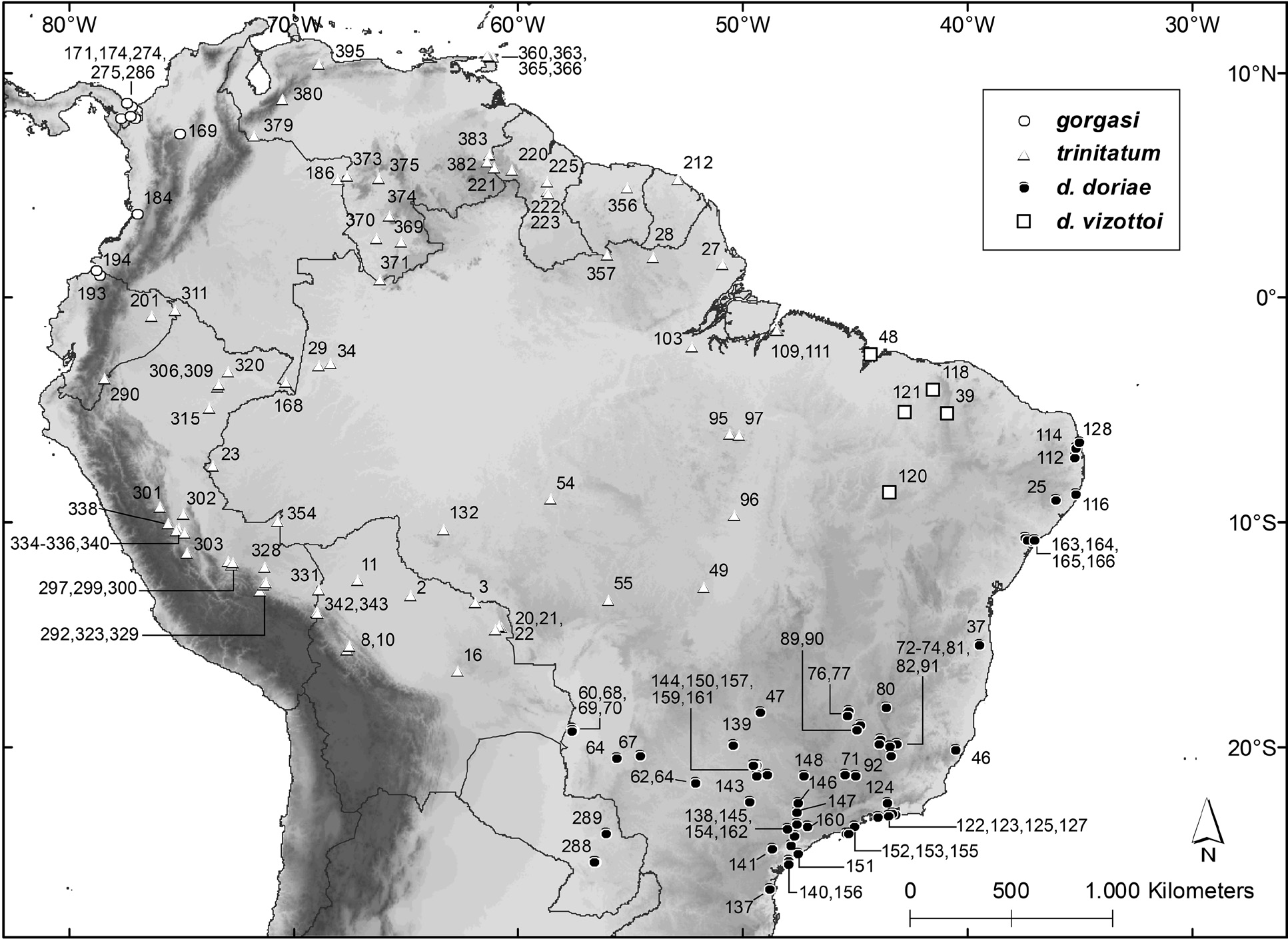
Distribution and Habitat. Specimens are known from Panamá, western Colombia, and northwestern Ecuador ( Fig. 23 View FIGURE 23 ), and there is a record from eastern Honduras ( Turcios-Casco et al. 2020). The unconfirmed record for Costa Rica is based on a bat captured and released by R. LaVal in Tortuguero ( Timm & LaVal 1998). Also in Costa Rica, Harvey & González-Villalobos (2007) reported the capture of 18 “ Chiroderma trinitatum ” in Talamanca , but we could not verify if there are voucher specimens to support this claim. The occurrence of the species in Costa Rica is expected, as C. gorgasi has been recorded in western Panamá and eastern Honduras ( Handley 1966b; Turcios- Casco et al. 2020).
The records of C. gorgasi are from the humid forests of the Chocó of Colombia, the Darién of Panamá, Caribbe- an lowland forests of Honduras, and montane forests of the Rio Magdalena valley in Colombia. The altitude where specimens have been obtained ranges from 30 m in Esmeraldas, Ecuador, to 975 m, in Tacarcuna, Panamá. There are Colombian records of the species occurring at 2,100 in Tolima and between 1,600 and 2,300 m in Risaralda ( Galindo-Espinosa et al. 2010; Castaño et al. 2018).
Description and Comparisons. Dorsal pelage may vary from pale to dark brown. Individual hairs of the dorsum are tricolored: the base is approximately ¼ of the hair length and dark brown, the middle band is approximately ½ of the length of the hair and varies from buff to pale gray, and the tip is about ¼ of the hair length and varies from pale to dark brown. Both pairs of facial stripes are conspicuous. The dorsal stripe is conspicuous in approximately half of the sample (47%, n=8); whereas, it is barely visible in nine specimens. The ear margins and base are paler than the remainder of the ear conch. The noseleaf has a simple tip, is brown in color, with pale margins on the horseshoe.
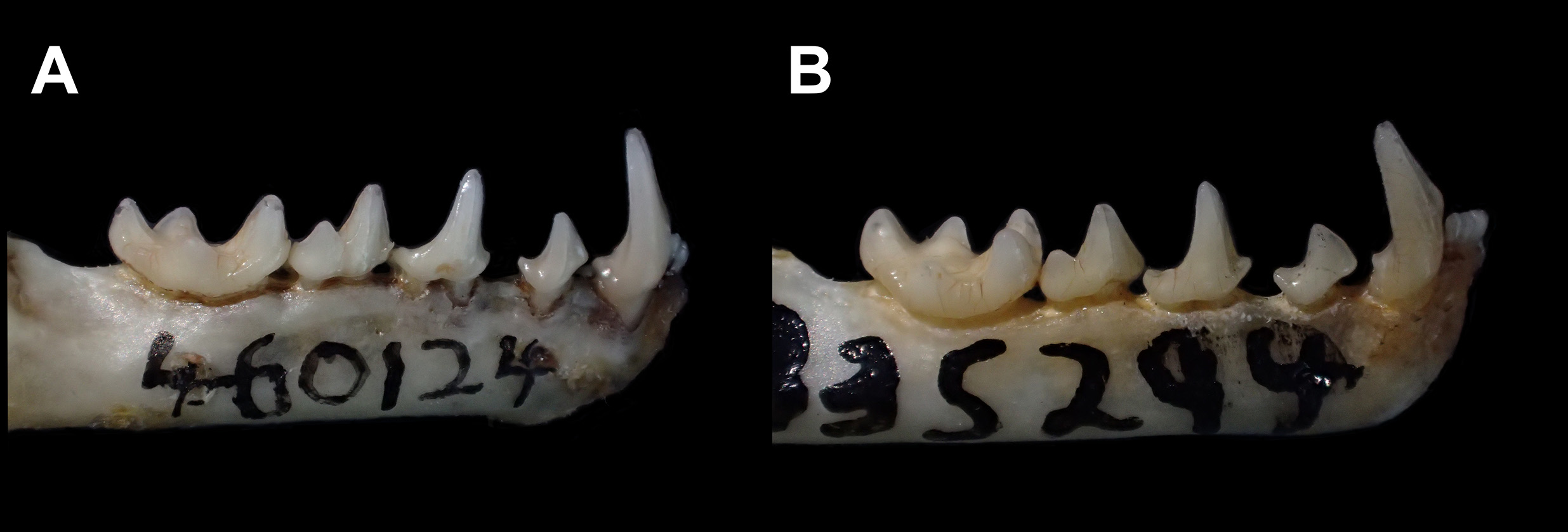
Dimensions of the skull are similar to those of C. trinitatum , and the two species are the smallest Chiroderma ( Tables 7 and 8). Braincase is globose, clearly distinguishable in profile from the frontonasal region. The sagittal crest is poorly developed and was not detected in 9 of the 17 specimens we scored for this character. The nasal notch is short and either does not reach the interorbital region, or extends only the level of the anterior border of the orbit. Similar to C. trinitatum , the post-orbital processes are rhomboid and not pointed as in the other Chiroderma . The posterior palatine process was absent in 13 of 15 specimens and in the other 2, the process was only a small bump. When cranium and mandible are in occlusion, a lateral gap is visible, bordered by C, P3, P4, p2 and p4.
The I1s are convergent and their tips are usually in contact. The mandibular condyle is level with or slightly below the toothrow. The lower canines are relatively narrow and high-crowned, with the crown tip level with the top of the coronoid process, when viewed laterally. The p2 is in contact with c, but not with p4, or if not in contact with the lower canine, p2 may be either closer to the lower canine or equidistant from c and p4. The p2 usually is longer mesiodistally than high and the protoconid is shifted anteriorly, not aligned with the base of the tooth when viewed laterally ( Fig. 27 View FIGURE 27 ).
Chiroderma gorgasi differs from every other Chiroderma , except C. trinitatum , by its smaller size and nasal notch usually not reaching the interorbital region. Comparisons with C. trinitatum were made in the previous section.
Geographic Variation and Phylogeography. Sequences of three individuals of C. gorgasi were analyzed in the phylogeny, precluding making inferences on geographical structuring. Within-species variation was 1.04%, the second highest value in Chiroderma after C. villosum (1.17%).
Subspecies. C. gorgasi is monotypic.
Remarks. Handley (1960) described Chiroderma gorgasi based on five specimens from Panamá and one C. trinitatum from Trinidad, the type and only known specimen at the time. In the original description, Handley (1960: 465) suggested that, as the sample size increased, the two taxa could prove to be conspecific. Shortly after its description, C. gorgasi was treated as a subspecies of C. trinitatum , based on their morphological similarity (Barriga- Bonilla 1965; Jones & Carter 1976; Hall 1981). Simmons (2005) recognized a monotypic trinitatum with gorgasi as a junior synonym. Recently, Lim et al. (2020) recognized C. gorgasi as a distinct species, because it does not share a most recent common ancestor with C. trinitatum , and has distinguishing morphological characters.
Natural History. C. gorgasi is a frugivore, specialized on fruits of Ficus ( Bonaccorso 1979) . Four species of fruits and infructescences have been recorded in the diet of C. gorgasi : Ficus popenoei , Piper aduncum , Solanum umbellatum , and Vismia sp. ( Bonaccorso 1979; Castaño et al. 2018). The vertical distribution suggests that C. gorgasi is a canopy and sub-canopy frugivore, more frequently captured in nets between 3 and 12 m above ground ( Bonaccorso 1979).
The few reproductive data for the species suggest a pattern of seasonal polyestry. A pregnant female was captured in June in Colombia and lactating individuals were recorded in February and March in Panamá. Literature data for Panamá report pregnancies in February, May and between September and November; whereas lactating females are documented from May and September ( Fleming 1973; Bonaccorso 1979). Births apparently occur toward the end of the dry season, between February and May, and in the middle of the rainy season, between July and September, when fruits are most abundant.
Specimens Examined (N = 18): Colombia: Antioquia, La Tirana ( IAvH-M 917 , 934 , 974 , USNM 499475 About USNM , 499477 About USNM , 499479 About USNM ) ; Chocó, Corregimiento Gilgal ( IAvH-M 4932 ), Finca El Recurso ( IAvH-M 3260 , 3299 , 3323 ) ; Valle del Cauca, Río Zabaletas ( USNM 483764 About USNM ) . Panamá: Darién, Parque Nacional Darién ( ROM 104342 About ROM ) , Tacarcuna Village Camp ( USNM 309902 About USNM , 309903 About USNM [holotype of gorgasi ], 309904 ) ; San Blas, Armila ( USNM 335294 About USNM , 335296 About USNM , 335297 About USNM ) .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Stenodermatinae |
|
Tribe |
Stenodermatini |
|
SubTribe |
Vampyressina |
|
Genus |
Chiroderma gorgasi Handley, 1960
| Garbino, Guilherme S. T., Lim, Burton K. & Tavares, Valéria Da C. 2020 |
Chiroderma trinitatum gorgasi : Barriga-Bonilla, 1965: 246
| Barriga-Bonilla, E. 1965: 246 |
Chiroderma gorgasi
| Handley Jr., C. O. 1960: 464 |