
Euura acutiserra ( Lindqvist, 1949 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4302.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:31B4D326-8D50-41A9-A8A7-69D4427BAD53 |
|
DOI |
https://doi.org/10.5281/zenodo.4902007 |
|
persistent identifier |
https://treatment.plazi.org/id/03B9953B-5C60-5911-FF48-F94C2014FCBC |
|
treatment provided by |
Plazi |
|
scientific name |
Euura acutiserra ( Lindqvist, 1949 ) |
| status |
|
Euura acutiserra ( Lindqvist, 1949)
Pontania acutiserra Lindqvist, 1949: 66 –68. Described: ♀, ♂. Holotype, ♀, FMNH [examined]. Type locality: Finland, Utsjoki , Outakoski.
Phyllocolpa acutiserra: Benson (1960a) .
Pontania (Phyllocolpa) acutiserra: Viitasaari & Vikberg (1985) .
Nematus (Phyllocolpa) acutiserra: Zhelochovtsev (1988) .
Euura acutiserra: Prous et al. (2014) .
Variability. Female: Body length: 3.7–4.0mm. Male: 3.6–4.0mm. Female and male: no significant variability in colour pattern. Total number of specimens examined: 17.
Genetic data. COI barcodes in BOLD of four E. acutiserra specimens from Norway and Finland with approx. 0.2% intraspecific variability diverge by a minimum of about 3.0% from the morphologically most similar north European species, E. leucapsis . The European E. acutiserra barcodes nest within a BIN containing sequences of seven Canadian specimens under the name Phyllocolpa excavata : the most similar of these sequences to a European specimen differs by about 1.0%.
Bionomics. Host plants: Salix lapponum ( Kopelke 2007b) ,? S. glauca ( Vikberg 1970, Zinovjev 1999). The record from S. glauca in Vikberg (1970) is based on a single specimen reared by E. Peltonen. Possibly the Salix species was misidentified, or S. glauca is a rarely used host. Males of E. acutiserra are rare ( Vikberg 1970, Kopelke 2007b): apart from the single male paratype, we examined two males reared by Kopelke. Biology: Kopelke (2007b), Zinovjev & Vikberg (1998).
Distribution. North Europe ( Kopelke 2007a), possibly including Scotland ( Benson 1958; but the identity of the single specimen needs checking, because neither recorded host is known at its collection site), Russia (Yakutia: Popov 2011). Recorded in Canada ( Benson 1962), but determination requires checking; see also above, under Genetic data. Occurrence in Sweden: published records; Jämtland (Haris 2009), Västerbotten ( Kopelke 2007b), Lycksele Lappmark, Torne Lappmark (Haris 2009). Material examined: Hälsingland, Jämtland, Lule Lappmark, Norrbotten, Torne Lappmark.
Euura anglica (Cameron, 1877) comb. nov.
Nematus anglicus Cameron, 1877a: 173 . Described: ♀. Lectotype, ♀, designated by Kopelke (2007a), BMNH [not examined]. Type locality: [?] England.
Phyllocolpa anglica (Cameron, 1877) : Benson (1960a), Vikberg (2010a).
Nematus (Phyllocolpa) anglicus: Zhelochovtsev (1988) .
Pontania (Phyllocolpa) anglica: Zinovjev (1993b) .
Nematus nigrolineatus Cameron, 1879 [mandatory correction of incorrect original spelling nigro-lineatus]: 108–110. Described: ♀, ♂, gall, recorded host: Salix viminalis . Lectotype, ♀, designated by Kopelke (2007a), BMNH [not examined]. Type locality: England, Worcester. Synonymy with anglica by Benson (1940b).
Pontania fibulata Konow, 1901a: 84 . Described: ♀, ♂. Lectotype, ♀, designated by Kopelke (2007a), SDEI [examined]. Type locality: Czech Republic, Moravia. Synonymy with anglica by Vikberg (2010a).
Phyllocolpa scotaspis: Kopelke 2007c ; misidentification.
Notes on types and taxonomy. See remarks under E. scotaspis .
Variability. Female: Body length: 3.6–3.9mm. Male: 3.5–3.8mm. Female and male: supraclypeal area and lateral vertex pale marked to completely black. Total number of specimens examined: 12.
Genetic data. A COI barcode for a single specimen from the Netherlands is nearly identical to sequences of E. leucapsis .
Similar species. External morphology and colour pattern resembles E. leucapsis .
Bionomics. Host plants: Salix viminalis , S. dasyclados ( Vikberg 2010a) . Biology: Benander (1969)
Distribution. Central Europe and British Isles (Taeger et al. 2006), southern parts of Scandinavia ( Benander 1969), Russian Far East ( Zhelochovtsev & Zinovjev 1995). Occurrence in Sweden: published records; Skåne ( Benander 1969, Coulianos & Holmåsen 1991).
Euura destricta ( MacGillivray, 1923) comb. nov.
Pontania destricta MacGillivray, 1923: 168 –169. Described: ♀. Lectotype, ♀, INHS [examined], designated by Frison (1927). Type locality: U.S.A., Alaska, Katmai.
Pontania apicifrons Malaise, 1931b: 33 –34. Described: ♀, ♂. Lectotype, ♀, designated by Kopelke (2007a), NHRS [ NHRS- HEVA 000005013 ; examined]. Type locality: Russia, Kamtchatka.
Pontania apicifrons var. punctifrons Malaise, 1931b: 34 . Described: sex not stated. Lectotype, ♀, designated by Kopelke (2007a), NHRS [NHRS-HEVA000005014; examined]. Type locality: Russia, Kamtchatka. According to Taeger et al. (2010) an infrasubspecific name and therefore unavailable. Name preoccupied in Euura by Pachynematus punctifrons Malaise, 1921 ( Malaise 1921b) .
Pontania carinifrons Benson, 1940b: 209 –211. Described: ♀, ♂, recorded host: Salix pentandra . Holotype, ♀, BMNH [examined]. Type locality: Scotland, Roxburghshire , Newcastleton. Syn. nov.
Phyllocolpa carinifrons: Taeger et al. (2010) .
Pontania excavata: Coulianos & Holmåsen (1991) ; misidentification.
Phyllocolpa excavata: Benson (1960a) ; misidentification.
Pontania (Phyllocolpa) excavata: Viitasaari & Vikberg (1985) ; misidentification.
Nematus (Phyllocolpa) excavatus: Zhelochovtsev (1988) ; misidentification.
Phyllocolpa plicaglauca Kopelke, 2007a: 99 –100. Described: ♀, ♂, gall, recorded hosts: Salix glauca and glaucosericea. Holotype, ♀, SMF [examined]. Type locality: Norway, Finnmark, N.- Varanger , Reppen. Syn. nov.
Notes on types and taxonomy. P. destricta . MacGillivray (1923) stated neither the number of specimens in the type series, nor made any reference to a specimen as the primary type. Zinovjev & Smith (2000) wrongly referred to the single extant specimen in INHS as the holotype.
Other North American specimens examined: U.S.A.: Alaska, 1♀, Sealer's Is., 10.07.1965, D. M. DeLong, USNM . Oregon, 1♂, Union Co., Red Bridge St. Pk. 10 mi. E Starkey, 26.05.1984, D. R. Smith, USNM ; 1♀, Deschutes Co., Tumalo St. Pk. 2 mi. W Tumalo, 0 4.06.1984, D. R. Smith, USNM . California, 1♀, Placer Co., 1 mi W of Baxter , 3800', 0 8.06.1980, L. G. Bezark, USNM . New York, 1♀, Franklin Co., 5mi. E of Tupper Lake , Malaise Trap, 0 9.06.1983, D. R. Smith, USNM . Canada: British Columbia, 1♀, Rogers Pass, creek margin, Glacier N. Park, 30.06.1968, W. W. Wirth, USNM . Alberta, 1♀, 17 mi. S Seebe, Kananaskis For. Exp. Sta., 04– 06.06.1978, D. R. Smith, USNM . Quebec, 1♀, James Bay Hwy km 221, sweeping, 06– 08.06.1985, H. Goulet & D. R. Smith, USNM ; 1♀, James Bay Hwy km 335, Malaise trap, 08– 19.06.1985, H. Goulet & D. R. Smith, USNM .
P. apicifrons . Additional type material examined: Paralectotypes 3♀, 3♂, NHRS.
P. plicaglauca . Kopelke (2007a) stated that "The morphological characters of the new species plicaglauca do not correspond with any other species of this group". The conspicuous carina along the antennal hollows, in our opinion unique to a single taxon amongst the European gall-makers, is not mentioned. Nor is this described for the segregate that he called carinifrons . His illustrations of the lancets and penis valves of carinifrons and plicaglauca show these to be practically identical.
The very similar morphology and coloration of the nominal taxa destricta , apicifrons , carinifrons and plicaglauca , combined with the congruence of COI sequence data for these, have led us to conclude that they are conspecific. The species has already been considered by some authors to have a Holarctic distribution (e.g. Benson 1958, Smith 1979), and the names destricta (type locality: Katmai, Alaska) and excavata (type locality: Colorado) have sometimes been used for European populations. Zinovjev & Smith (2000) doubted whether excavata and destricta are conspecific. Kopelke (2007a) rejected the hypothesis that carinifrons is conspecific with any of the Nearctic taxa, arguing that carinifrons cannot occur in N. America, where Salix pentandra grows only as an introduced species. However, since carinifrons seems to have more than one host, and one of these, S. glauca , is widespread in North America, this argumentation is not convincing. After examining all relevant types, we agree with Zinovjev & Smith (2000), that excavata and destricta are not conspecific. See below under E. megacephala for further discussion.
Variability. Female: Body length: 4.1–4.9mm. The apex of valvula 3 in lateral view is usually quite obtuse, but not infrequently more acute, both in specimens reared from S. pentandra and S. glauca . Male: 3.7–4.4mm. Female and male: The colour pattern and morphology of this species, including surface sculpture, are remarkably stable throughout most of its range. The primary types of destricta , apicifrons and carinifrons differ only slightly in the extent of pale on the femora, pronotum, and lateral vertex. Total number of specimens examined: 52.
Genetic data. BOLD barcode data for 21 European and Canadian specimens under the names Phyllocolpa excavata and P. carinifrons share the same BIN. Intraspecific variability within this BIN is approximately 1.6%, attributable largely to the Canadian specimens (Alberta, Manitoba, British Columbia). Four specimens from Finland ( GenBank accession EU083959 View Materials ), Sweden, Norrbotten (DEI-GISHym20679), Scotland (DEI- GISHym5032) and Canada, Manitoba (10PROBE-21909) have an identical barcode. The nearest neighbouring European species, with a minimum of 2% difference, is E. leucapsis .
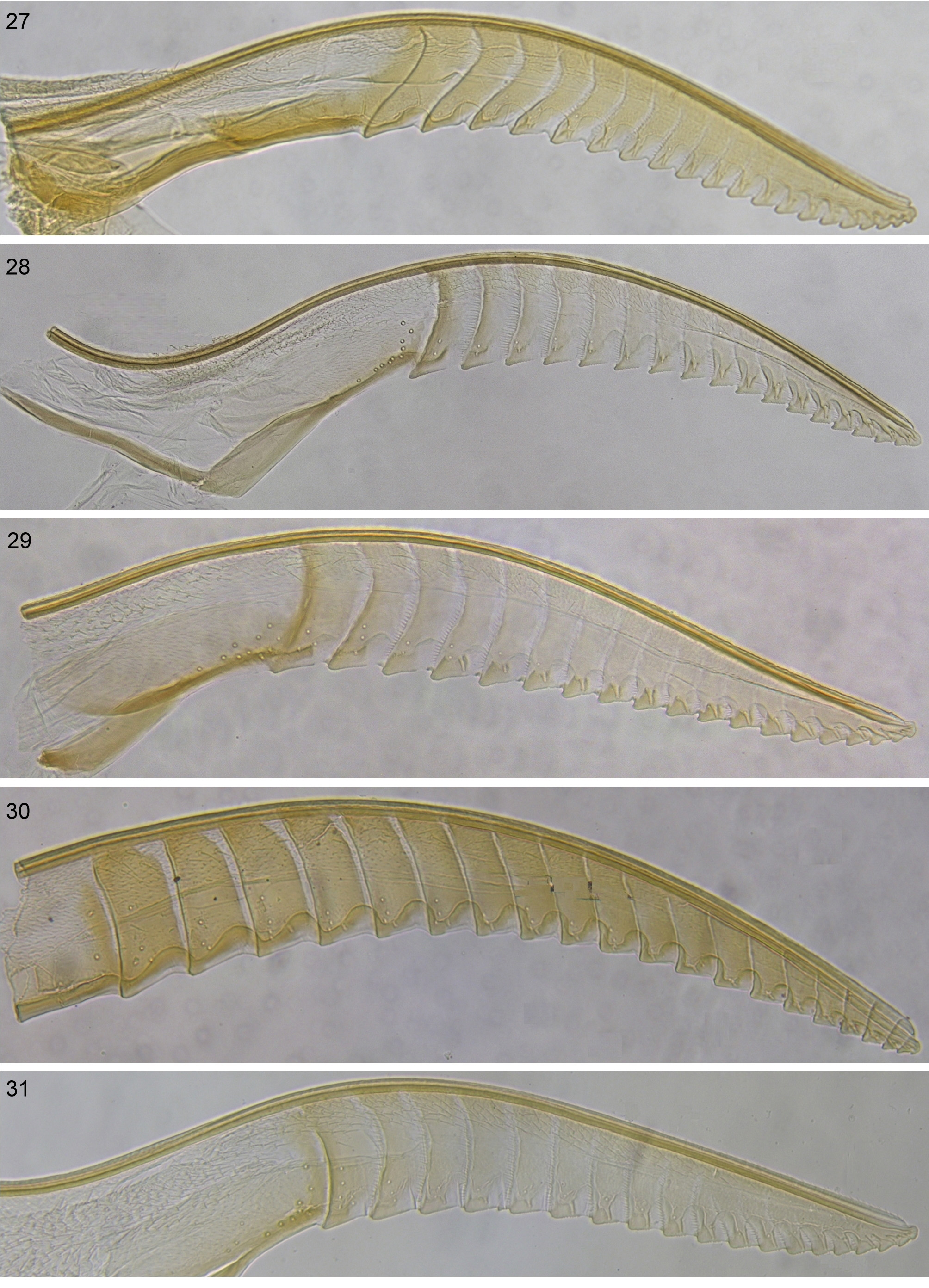
Similar species. Morphologically, including their lancets, E. destricta strongly resembles species of the ischnocera complex. The latter lack the carina along the antennal hollows, and annular suture 2 has a fully developed band of ctenidia: Fig. 28 View FIGURES 27 – 31 (ctenidia very few, or absent, on annular suture 2 in E. destricta : Fig. 29 View FIGURES 27 – 31 ).
Bionomics. Host plants: main hosts are Salix pentandra ( Kopelke 2007a, as Phyllocolpa carinfrons ), Salix phylicifolia ( Vikberg 1970, as Pontania excavata ), and Salix glauca ( Kopelke 2007a, as P. plicaglauca ). S. myrtilloides and S. lapponum are probably additional, perhaps infrequently used hosts (below). Kopelke (2007a) rejected Vikberg's record of S. phylicifolia as a host at Kilpisjärvi, northern Finnish Lapland, because he held P. carinifrons to be strictly monophagous on S. pentandra . Females from Kilpisjärvi, reared from S. phylicifolia , are morphologically indistinguishable from specimens reared from S. pentandra , S. glauca and S. lapponum . Furthermore, several E. destricta were collected by sweeping at subalpine levels near Abisko, Swedish Lapland, at a site where no S. pentandra occurs. Here, S. phylicifolia , S. glauca and /or S. lapponum may be the hosts. That S. myrtilloides is also a host is indicated by sequence data, identical to samples from S. pentandra and S. glauca , for a larva ( AS _ Euura _species_ Salix _ myrtilloides , leg. Nyman) collected in Finland from this host Three females and five males reared by Kopelke from S. lapponum and determined by him partly as P. acutiserra , partly as P. plicalapponum , are morphologically indistinguishable from destricta reared from other hosts. These specimens are all from a single locality: Norway, S. Trøndelag, Meldal. In view of this, the possibility of misidentification of the willow should be borne in mind, and use of S. lapponum as a host by E. destricta therefore requires confirmation. Biology: Kopelke (2007a).
Distribution. Central and North Europe including Britain and Ireland (Taeger et al. 2006), north to Kilpisjärvi in Finland; E. Siberia and Russian Far East ( Zhelochovtsev & Zinovjev 1995); N. America (northern Canada, and U.S.A., south to California, in the Sierra Nevada Mountains). Occurrence in Sweden: published records; Skåne ( Benander 1969), Halland ( Andersson 1955: as P.? leucaspis [sic!] on S. pentandra ), Jämtland, Västerbotten (Haris 2009), Torne Lappmark ( Coulianos & Holmåsen 1991). Material examined: Ångermanland, Västerbotten, Torne Lappmark.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Euura acutiserra ( Lindqvist, 1949 )
| Liston, Andrew D., Heibo, Erik, Prous, Marko, Vårdal, Hege, Nyman, Tommi & Vikberg, Veli 2017 |
Phyllocolpa plicaglauca
| Kopelke 2007: 99 |
Pontania acutiserra
| Lindqvist 1949: 66 |
Pontania carinifrons
| Benson 1940: 209 |
Pontania apicifrons
| Malaise 1931: 33 |
Pontania apicifrons var. punctifrons
| Malaise 1931: 34 |
Pontania destricta
| MacGillivray 1923: 168 |
Pontania fibulata
| Konow 1901: 84 |
Nematus anglicus
| Cameron 1877: 173 |