
Dryobalanoxylon sp.
|
publication ID |
https://doi.org/10.5252/geodiversitas2022v44a28 |
|
publication LSID |
urn:lsid:zoobank.org:pub:2611B0BC-F569-4135-A09C-6E527C2565A4 |
|
DOI |
https://doi.org/10.5281/zenodo.7157609 |
|
persistent identifier |
https://treatment.plazi.org/id/03B787F6-A21D-FFAF-893F-FF0AE6A7E8D3 |
|
treatment provided by |
Felipe |
|
scientific name |
Dryobalanoxylon sp. |
| status |
|
( Fig. 14 View FIG )
MATERIAL. — MNHN.F.50191 (field number: 17FN09). Estimated minimal diameter: 10-22 cm.
LOCALITY. — Kalewa Township, Sagaing Region, Myanmar.
AGE. — Upper lower to lowermost middle Miocene.
DESCRIPTION
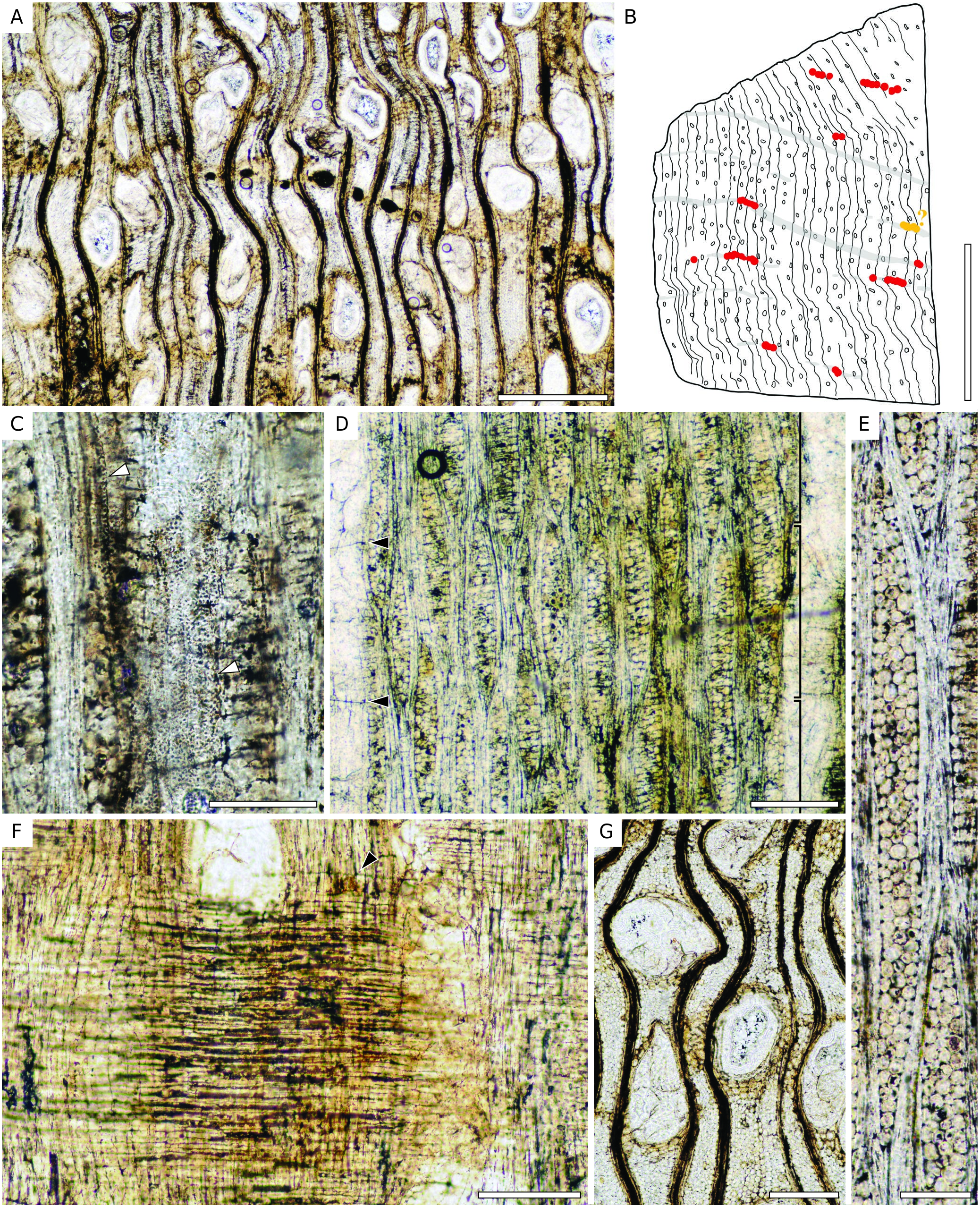
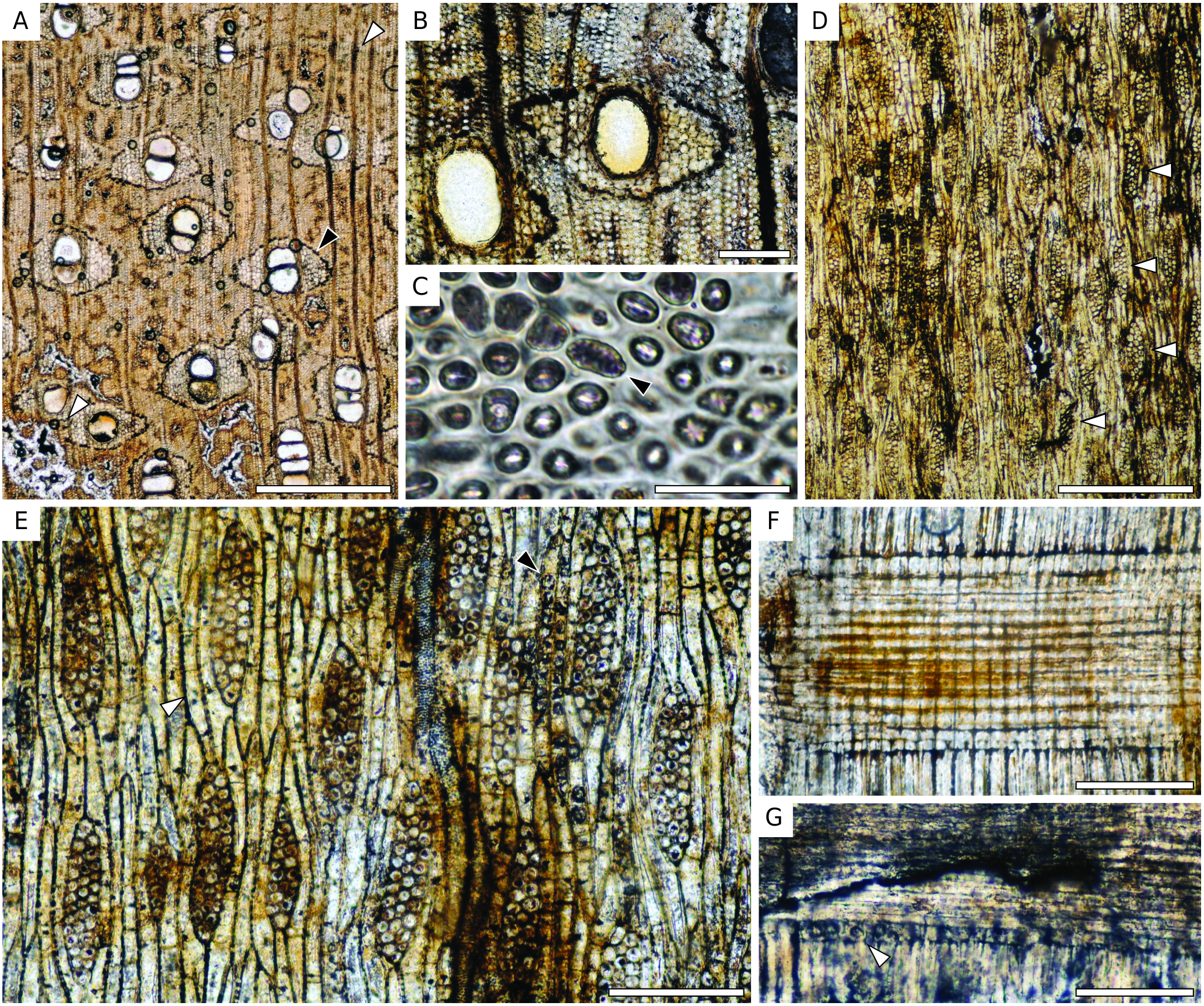
Wood diffuse-porous. Growth rings indistinct or absent. Vessels mostly solitary (81-90%) ( Fig. 14A View FIG ) or in radial groups of 2(-3), round to oval, 5-13 per mm² (average: 8); tangential diameter 140-260 µm (average: 200 µm). Tyloses present ( Fig. 14G View FIG ). Vessel elements 140-290 µm (average: 210 µm) long. Perforation plates simple ( Fig. 14D View FIG ). Intervessel pits alternate, 3-6 µm of diameter (average: 5 µm). Vessel-ray pits not preserved. Vasicentric tracheids present ( Fig. 14C View FIG ). Parenchyma aliform with small wings, or at least vasicentric forming a sheath of 1-3 cells around vessels ( Fig. 14A, G View FIG ); occasionnaly confluent; diffuse parenchyma may occur but not as an obvious feature. The parenchyma also seems to be in tangential bands, 1-6 seriate, possibly crossing the whole section or at least some portion; they can contain secretory canals ( Fig. 14A, B View FIG ); these bands are separated by 1.5-4 mm. Parenchyma cells 75-130 µm long (average:110 µm) 10-30 µm wide (average: 20 µm) in tangential section; without crystals. Rays 1- to 4-seriate, mostly 3-4-seriate ( Fig. 14D, E View FIG ), with a possible storied tendency, but faint and limited to zones with shorter rays ( Fig. 14D View FIG ), 6-9 rays per mm (average: 8), 320- 1140 µm (average: 540 µm) or 15-50 cells high, heterocellular with 1-(maybe more?) upright or square cell at the ends to possibly homocellular ( Fig. 14F View FIG ); sheath cells absent or very rare, end-to-end fusion present but not frequent. Fibres nonseptate, 9-21 µm (average: 15 µm) wide, walls commonly not preserved. Secretory canals in short (to long) lines of 2-5 (-7) canals surrounded by parenchyma ( Fig. 14A View FIG ), 40-90 µm of diameter (average: 60 µm). These lines seem to be parts of longer bands of parenchyma, but the compression of the wood may have crushed canals, which do not allow us to determine if lines are longer. Some scattered canals found.
DISCUSSION
This specimen has: 1) vertical secretory canals in short to long tangential lines or diffuse; 2) mostly solitary vessels; 3) vasicentric tracheids; 4) up to more than 1 mm long and mainly 3-(4-)seriate rays ( Fig. 8D, E View FIG ); and 5) tyloses. Vasicentric tracheids and axial secretory canals indicate an affinity with the Dipterocarpaceae family. Features of the specimen could fit the diagnosis of both Dipterocarpus and Dryobalanops genera. This greatly depends on author’s definitions of “short tangential lines” ( Metcalfe & Chalk 1950; Schweitzer 1958; IAWA Committee 1989). Schweitzer (1958) highlighted the fact that the inter- and intraspecific variation are sometimes too wide to define precise limits between these genera. Solitary secretory canals together with lines up to 7 canals, the parenchyma pattern (vasicentric and aliform), the size of the vessels, the relatively thin rays (up to 4-seriate) recall Dipterocarpus ( Metcalfe & Chalk 1950; Gottwald & Parameswaran 1966; Soerianegara & Lemmens 1993; Ogata et al. 2008). The presence of tylose, of weakly heterocellular rays (mostly one row of upright marginal cells), quite thin and short rays, the quasi-absence of sheath cells, and very few uniseriate recall Dryobalanops even though the unilateral arrangement of the parenchyma is not obvious ( Metcalfe & Chalk 1950; Gottwald & Parameswaran 1966; Soerianegara & Lemmens 1993; Ogata et al. 2008). The canals we identified are relatively small (<100 µm) and they seem to occur in tangential bands of parenchyma. These bands cross at least a part of the section and it is possible that they contain two of these lines of secretory canals. As they are relatively regularly spaced, they could be interpreted as long tangential band of secretory canals, if we consider that most of them are crushed by the compression of the wood ( Fig. 14B View FIG ). After a comparison with modern species of both genera, we note a close affinity with Dryobalanops aromatica C.F.Gaertn. (specimens CTFT19577 and MNHN-P-P00402611) that displays frequent vessels, slightly aliform parenchyma as well as diffuse, no silica bodies in ray cells, rays 2-4-seriate with numerous 3-seriate, fusiform rays with short rows of marginal cells (1 or 2 cells) that are sometimes storied, and similar thin tangential bands of parenchyma that can contain diffuse to long lines of canals interrupted by vessels. Dryobalanops aromatica differs from our fossil in the absence of tylose and the presence of diffuse-in-aggregate parenchyma. Dryobalanops oblongifolia from InsideWood (2004 -onward) displays regular tangential lines of secretory canals embedded in parenchyma bands as well as a pattern of parenchyma that matches the one of our fossil, but its rays are broader (up to 7-seriate in the pictures provided). Due to the strong resemblance and variation among Dryobalanops species, we only attribute as NLR of this fossil the genus Dryobalanops , with a noted affinity for D. aromatica .
The fossil genus Dryobalanoxylon was instituted for fossil woods resembling Dryobalanops ( Den Berger 1923; Schweitzer 1958). The only difference within the diagnosis of Dryobalanoxylon and Dipterocarpoxylon (extracted from Schweitzer 1958) is that Dryobalanoxylon can have storied structures, somehow smaller vessels, secretory canals mostly in long tangential bands, rarely short, and common fibre-tracheids. No distinction is made based on rays; however, it can be seen within the described species, more or less the same differences as between modern species. Although we do not observe any fibre-tracheids, probably because of the poorly preserved cell walls, an examination of the resembling species shows a closer affinity for Dryobalanoxylon ( Den Berger 1927; Schweitzer 1958; Awasthi 1971; Vozenin-Serra 1981; Awasthi & Ahuja 1982; Mandang & Kagemori 2004; Biswas et al. 2019) especially: D. sumatrense Schweitzer , D. tobleri Den Berger , D. lunaris Mandang & Kagemori , D. holdeniae (‘ holdenii ’) Awasthi and D. tambouense Vozenin-Serra ( Appendix 1). Few differences distinguish these Dryobalanoxylon species from each other as the descriptions display a great degree of overlapping. We note a closer resemblance with D. tambouense , which has vasicentric to aliform parenchyma, a similar size of rays (2-5-seriate, up to 1194 µm) with a storied tendency, and a similar size and frequency of vessels. The state of preservation of our fossil and its unclear canal distribution pattern does not allow us to attribute the specimen to an individual species with certainty nor to create a new species. Thus, we name this fossil Dryobalanoxylon sp. Dryobalanops general ecology and distribution is givenp. 878; Dryobalanops aromatica lives in lowland mixed dipterocarp forests, on dry sandy or gravelly soils near the coast up to 400 m ( Ashton 1982; Tropical Plant Database 2014 -onward; Barstow & Randi 2018). Dryobalanops oblongifolia lives in lowland mixed dipterocarp forests, also in periodically inundated, freshwater swamps, near streams, in poorly drained forests or on hillsides below 600 m ( Ashton 1982; Soerianegara & Lemmens 1993; Barstow 2018a).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |