
Tarphionivea lindsayae Leschen and Lord, 2017
|
publication ID |
https://doi.org/10.1649/0010-065X-71.4.733 |
|
publication LSID |
urn:lsid:zoobank.org:pub:FDCD5A33-AC6A-4BF1-A833-208A371861EA |
|
persistent identifier |
https://treatment.plazi.org/id/D787E18D-05B0-4D99-B2D4-F8E68B8C0815 |
|
taxon LSID |
lsid:zoobank.org:act:D787E18D-05B0-4D99-B2D4-F8E68B8C0815 |
|
treatment provided by |
Diego |
|
scientific name |
Tarphionivea lindsayae Leschen and Lord |
| status |
sp. nov. |
Tarphionivea lindsayae Leschen and Lord , new species
( Figs. 2a–c View Fig , 3a–n View Fig )
Type Material. Holotype: male [missing left metatibia and tarsus, genitalia removed], “ NEW ZEALAND, NN, Mt Arthur Hut , 23 Jan 2012, R. Leschen, L. Dunning, H. Lindsay, 41.197494S, 172.715921E, beating beech trees with white lichen at night, RL1627” GoogleMaps . Paratypes: 2, same data as holotype; 1, AMNZ [missing left protarsus and left metatarsus]: “ Balloon Hut Mt Arthur Plateau 2-1-48 C. E. C. [handwritten in pencil]/ C. E. Clarke Collection ” GoogleMaps .
Description. Cuticle dark to light reddish brown; tarsi and mouthparts red-brown to light brown. Vestiture consisting of elongate scales that form agglutinate patches dorsally, especially dense on the genae, more or less evenly distributed ventrally; mostly golden and silver scales in patchworks on anterior portion of elytra and more or less white in small median spot and 1 subapical spot and a subapical transverse row on the subapical transverse ridge; scales of legs dark brown to black, tibial apices paler. Surfaces rugose; irregular punctures on surfaces distinct, forming weak semi-seriate rows on elytra, connected by channels interrupted by tubercles and carinae; ventral surfaces with distinct verrucae. Ratios among antennomere lengths: 2.3/ 3.0/2.5/2.0/2.4/1.1/1.2/1/1.1/2.0/2.5. Elytra with up to 5 basal costae including the humeral costa, outer subhumeral costa may be tuberculate and/or may fuse with the humeral (4 th) costa, which extends posteriorly to basal 1/4, terminating on a tubercle; with discal tubercles present at basal 1/3 and 2/3, with a large subapical tubercle at apical 1/4 that may be transverse and form a carina, with 2 smaller tubercles just in front of and laterad of large subapical tubercle. Aedeagus about 4X as long as wide, parallel-sided and slightly expanded apically; parameres slender, about as long as phallobase, each paramere somewhat flattened; penis narrow and approximately 1.3X as long as parameres, apex subacute, with thin subapical flanges, notched at base.
Comments. Tarphionivea lindsayae is known from four specimens, three of which were beaten at night from a dry, rotten Nothofagus tree high in the canopy level reached along the edge of a track above Mt. Arthur Hut and skirting the margin of the alpine tussock zone. The locality at Mt. Arthur is well known among entomologists and visited relatively frequently, indicating that the species may be a canopy specialist. Balloon Hut is at a slightly lower altitude than Mt. Arthur Hut but of similar habitat and located in tableland.
We recommend that T. lindsayae be placed on the Department of Conservation Threatened Species List ( Leschen et al. 2012) and treated as “rangerestricted” and “sparse” using the criteria of Stringer and Hitchmough (2012). This formality will facilitate further collecting activity to discover more about the biology and distribution of T. lindsayae .
Etymology. Patronymic for conservationist Helen Lindsay, in recognition of her leadership in habitat restoration throughout New Zealand and who was a participant in the fieldwork that led to the discovery of Tarphionivea .
Zebitoma Leschen and Lord , new genus ( Figs. 4a–i View Fig , 5a–m View Fig )
Type Species. Bitoma nana Sharp, 1876 ( Fig. 4e View Fig ).
Diagnosis. Surfaces scalate, agglutinate or not; with or without encrustations. Head not strongly dorsoventrally flattened; temples relatively long. Antenna 11-segmented with 2-segmented club. Antennal groove short and well-developed at anterior margin of eye. Labial palpi absent. Pronotum explanate or not posteriorly, anterior margin not deflected dorsally, lateral lobes present or not with anterior angles absent or present and not extending forward to level of anterior pronotal margin. Prothorax without distinct antennal cavities. Abdominal ventrites well separated by transverse grooves, ventrite V with or without well-developed preapical groove. Scutellary shield present and well-developed. Elytron with or without costae; epipleuron present to apex. Metacoxal cavity open laterally. Legs generally weakly flattened in cross-section. Tarsal formula 4-4-4; tarsi filiform, not strongly lobed below.
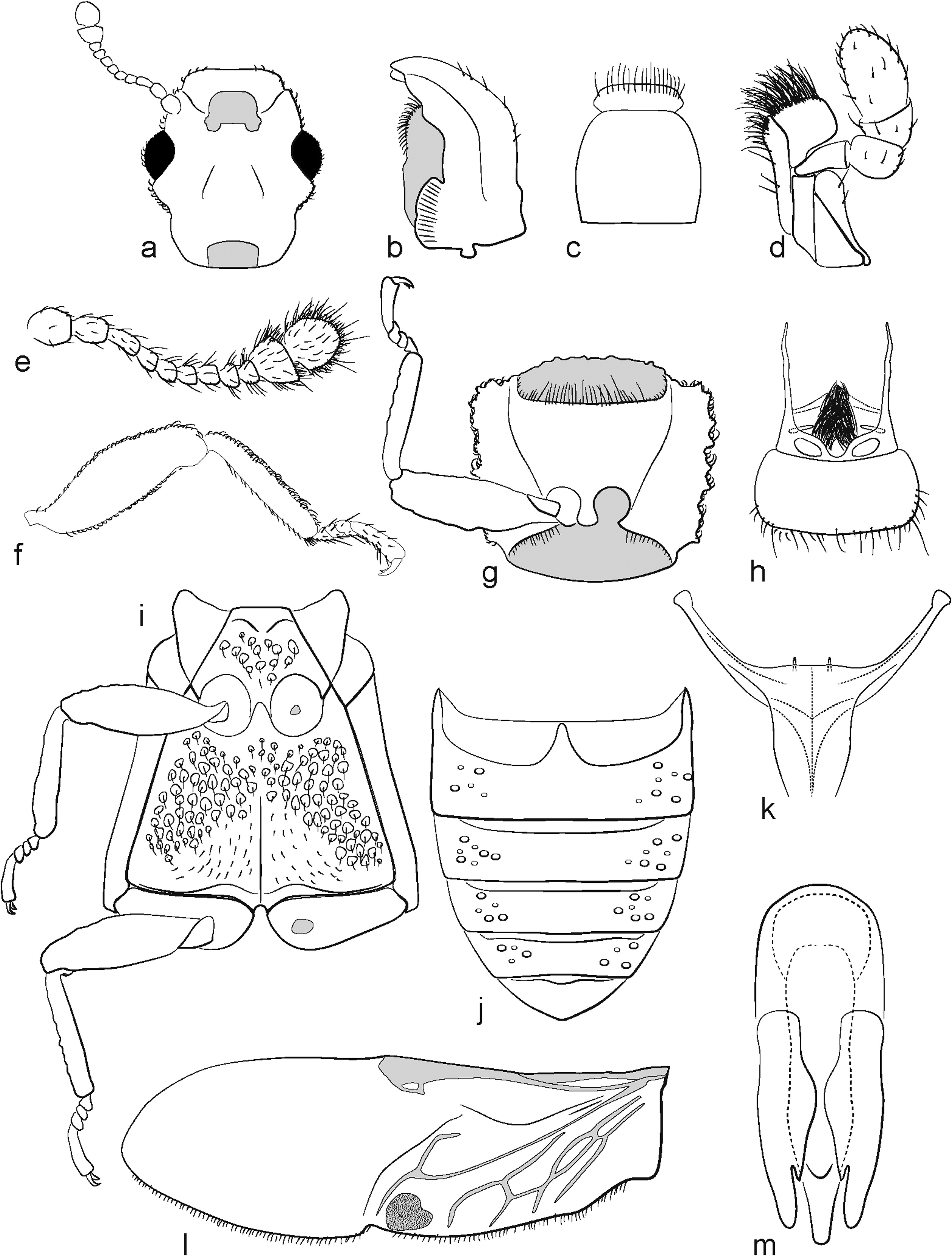
Description. Length 1.6–2.9 mm, greatest depth 0.5–1.1 mm. Dorsal and ventral surfaces scalate, often agglutinate, rugose-punctate, often tuberculate, with scales sometimes arising from tubercles; encrustations present or absent. Head not flattened, strongly constricted behind eyes ( Fig. 5a View Fig ); temples not strongly produced laterally, length, in dorsal view, equal to length of eye or slightly longer; frons typically depressed at middle, with sides distinctly or weakly raised above antennal insertions; supraorbital carina absent. Antennal groove well developed only at anterior margin of eye, never extending below; subgena not expanded laterally to form ventral shelf. Eyes well-developed, weakly protruding, convex, interfacetal setae present. Antenna short, inserted below frons, extending to level just beyond midline of pronotum, 11–segmented with 2-segmented club ( Fig. 5e View Fig ); setation of antennal segments 2–9 sparse, consisting of a single row of short, hair-like setae at base of segment; club segments more densely setose, not arranged in rows; scape subcylindrical, narrower than pedicel; pedicel not strongly constricted at base and weakly expanded distally or not, slightly curved, slightly longer than wide, shorter than antennomeres 1 and 3; antennomeres 3–9 of subequal width, becoming gradually shorter and weakly transverse; antennomere 3 distinctly longer than 4 or not; club not flattened, strong and abrupt, with antennomere 10 transverse and 11 subequal to 2X length of antennomere 9, well-separated. Labrum moderately long, quadrate, anterior margin slightly to moderately arcuate or more or less straight with apicolateral angles rounded, fringed with sparse setae ( Fig. 5h View Fig ). Mandibles symmetrical, bidentate, with welldeveloped hyaline prostheca; mola present and striate ( Fig. 5b View Fig ). Maxillary palpus 4-segmented, last maxillary palpomere apically truncate, about 2.5X as long as wide ( Fig. 5d View Fig ); galea narrowest at base, expanded apically, apical margin truncate, fringed with setae; lacinia narrower, with single, curved spine at apex, apical margin fringed with setae. Mentum transverse, without shallow, median impression, bearing a few short setae, anterior margin straight; labial palpi absent ( Fig. 5c View Fig ). Submentum somewhat trapezoidal, slightly wider than long; lacking a median carina. Gular region not broadly vaulted at middle, with visible sutures and tentorial pits. Prothorax constricted at base, with a narrow cowling fitting over the mesothorax. Pronotum about 1.2X wider than long, explanate (or not posteriorly in some species), anterior margin not deflected dorsally, lateral lobes present (up to three) or not, with anterior angles absent or present and do not extend forward to level of anterior pronotal margin ( Fig. 5g View Fig ); pronotum of most species widest at apex or apical 1/3, in most species narrowest at base with base narrower than combined width of elytra; anterior margin produced at middle in some species, but not forming a hood and not deflected dorsally at the anterior margin; posterior angles rounded to angulate, with or without a tooth; lateral carina sharp-edged in most species; surface of disc typically uneven, with weak depressions and elevations, sometimes with tubercles and distinctive costae. Prothoracic venter without distinct antennal cavities, though weakly excavate. Notosternal suture present. Prosternum not produced anteriorly into a chin-piece, anterior margin straight, medially not raised above level of procoxae; prosternum long in front of procoxae, about 2X as long as prosternal process; prosternal process narrower than coxal cavity, slightly expanded apically, apex subacute, weakly expanded apically or laterally, extending just beyond coxal cavities. Procoxal cavities externally separated by at most 1/3 coxal diameter, narrowly separated internally; externally narrowly open, postcoxal process acute and extending to about mid-length of exposed part of coxae. Lateral portions of elytra weakly to moderately explanate. Scutellary shield present, visible externally. Elytra 1.3X as long as combined width, 2.6X as long as pronotum; parallel-sided or weakly expanded towards apex; scutellary striole present, punctation sometimes obscured by rugose agglutinate surface; disc weakly or not tuberculate, sometimes with low costae that are more prominent apically or at humeral angles; apex subrounded in dorsal view; epipleuron more or less horizontal and visible slightly (especially anteriorly) in lateral view, complete to apex. Hindwings present ( Fig. 5l View Fig ). Mesoventrite ecarinate; mesocoxae very narrowly separated by less than 1/2 width of coxal cavity; mesocoxal cavity laterally closed ( Fig. 5i View Fig ). Metaventrite 1.5X as long as abdominal ventrite I; metacoxae separated by about 1/4 width of coxal cavity; metacoxal cavity laterally open ( Fig. 5i View Fig ). Legs generally weakly flattened in cross-section; outer apical angle of protibia not expanded; edge of protibia simple, without teeth or spines ( Fig. 5f View Fig ). Tarsal formula 4-4-4; tarsi filiform and not strongly lobed below, tarsomeres I–III subequal in length, their combined length subequal or shorter than tarsomere IV. Abdominal ventrites well separated by transverse grooves, intercoxal process of ventrite I acute ( Fig. 5j View Fig ); ventrite V with or without welldeveloped preapical groove, with apex abruptly, obliquely deflected ventrally in some species. Male sternite VIII with baculae well-separated, not meeting at midline; sternite IX with elongate and acute spiculum gastrale. Aedeagus with tegmen dorsal to penis ( Fig. 5m View Fig ).
Comments. The New Zealand members of Bitoma are highly variable. We have examined externally the relevant type material, but a majority of the species, including new ones, require full dissection and detailed morphological study. Zebitoma is described for nine species of New Zealand Bitoma that are easily separated from members of Bitoma sensu stricto by the absence of labial palpi and the anterior angles of the prothorax absent, or when present, the angles do not extend forward to the same level of anterior pronotal margin. Zebitoma can also be distinguished from New Zealand Bitoma sensu stricto by having a relatively smaller and subquadrate prothorax that is narrower than the combined width of the elytral bases and the presence of 1–3 lateral lobes in some species. Species of Zebitoma also tend to be smaller in size than members of Bitoma sensu stricto.
Apart from three species that we currently consider are valid members of Bitoma ( B. costicollis , B. insularis , and B. vicina ), B. mundula (and possibly the similar-looking B. auriculata ) may eventually be placed in Ablabus Broun based on the presence of a declivate elytral apex and lateral lobes on the pronotum, features present in the type species, Ablabus ornatus Broun and other members of Ablabus (preliminary molecular data also supports this placement of B. mundula in Ablabus ). These two species, and the remaining four species of Bitoma sensu stricto, lack the distinctive discal ridges on the pronotum as are present in the type species of the genus, Bitoma crenata (Fabricius) , and in B. costicallis and B. insularis (these latter two species are probably synonyms, compare color patterns in figs. 478 and 59 in Lord and Leschen 2014). Five species, B. auriculata , B. distincta , B. morosa , B. mundula , and B. serraticula , have explanate pronota, while B. auriculata also has a wide apical lobe reminiscent of some taxa placed in the polyphyletic genus Pristoderus . Lastly, B. distans has a relatively convex and narrow prothorax, resembling Zebitoma , but with a complete labium; its phylogenetic placement is likely derived from within Bitoma sensu stricto as sister taxon to B. insularis . Reliance on prothoracic characters alone is problematic for generic-level diagnostics. The absence of discal carinae, which is distinctive in many Bitoma species , also occurs in B. morosa and B. serraticula . Thus, more comprehensive morphological
investigations will be required to place these remaining species.
Members of Bitoma and Zebitoma are active at night, presumably saproxylic, and can be beaten from branches of various tree species or taken from fungus ( e.g., Kuschel 1990), under bark, or hand collected from the surfaces of standing trees. Detailed natural history information is sparse, and at least one montane species of Zebitoma is regularly collected at night on white, thin-layered lichen.
Etymology. The name combines the prefix “ Ze -” referring to New Zealand and the name Bitoma .
Included Species. Z. brouni (Hetschko, 1928) , new combination ( Fig. 4a View Fig ); Z. discoidea (Broun, 1880) , new combination ( Fig. 4b View Fig ); Z. guttata (Broun, 1886) , new combination ( Fig. 4c View Fig ); Z. lobata (Broun, 1886) , new combination ( Fig. 4d View Fig ); Z. nana (Sharp, 1876) , new combination ( Fig. 4e View Fig ); Z. novella (Hetschko, 1929) , new combination ( Fig. 4f View Fig ); Z. picicornis (Broun, 1909) , new combination ( Fig. 4g View Fig ); Z. rugosa (Sharp, 1876) , new combination ( Fig. 4h View Fig ); Z. scita (Broun, 1886) , new combination ( Fig. 4i View Fig ).



| R |
Departamento de Geologia, Universidad de Chile |
| L |
Nationaal Herbarium Nederland, Leiden University branch |
| H |
University of Helsinki |
| AMNZ |
Auckland Institute and Museum |
| C |
University of Copenhagen |
| E |
Royal Botanic Garden Edinburgh |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |