
Tylototriton ngarsuensis, Grismer & Wood Junior & Quah & Thura & Espinoza & Grismer & Murdoch & Lin, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4500.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:3518D551-CA4B-43A4-B6D3-8A159E2398E1 |
|
persistent identifier |
https://treatment.plazi.org/id/E6FE8B3C-56E9-4C07-B82E-D5F62FFA8B69 |
|
taxon LSID |
lsid:zoobank.org:act:E6FE8B3C-56E9-4C07-B82E-D5F62FFA8B69 |
|
treatment provided by |
Felipe |
|
scientific name |
Tylototriton ngarsuensis |
| status |
sp. nov. |
Tylototriton ngarsuensis View in CoL sp. nov.
Suggested common name: Ywangan Crocodile Newt
Figs. 4 View FIGURE 4 , 5 View FIGURE 5
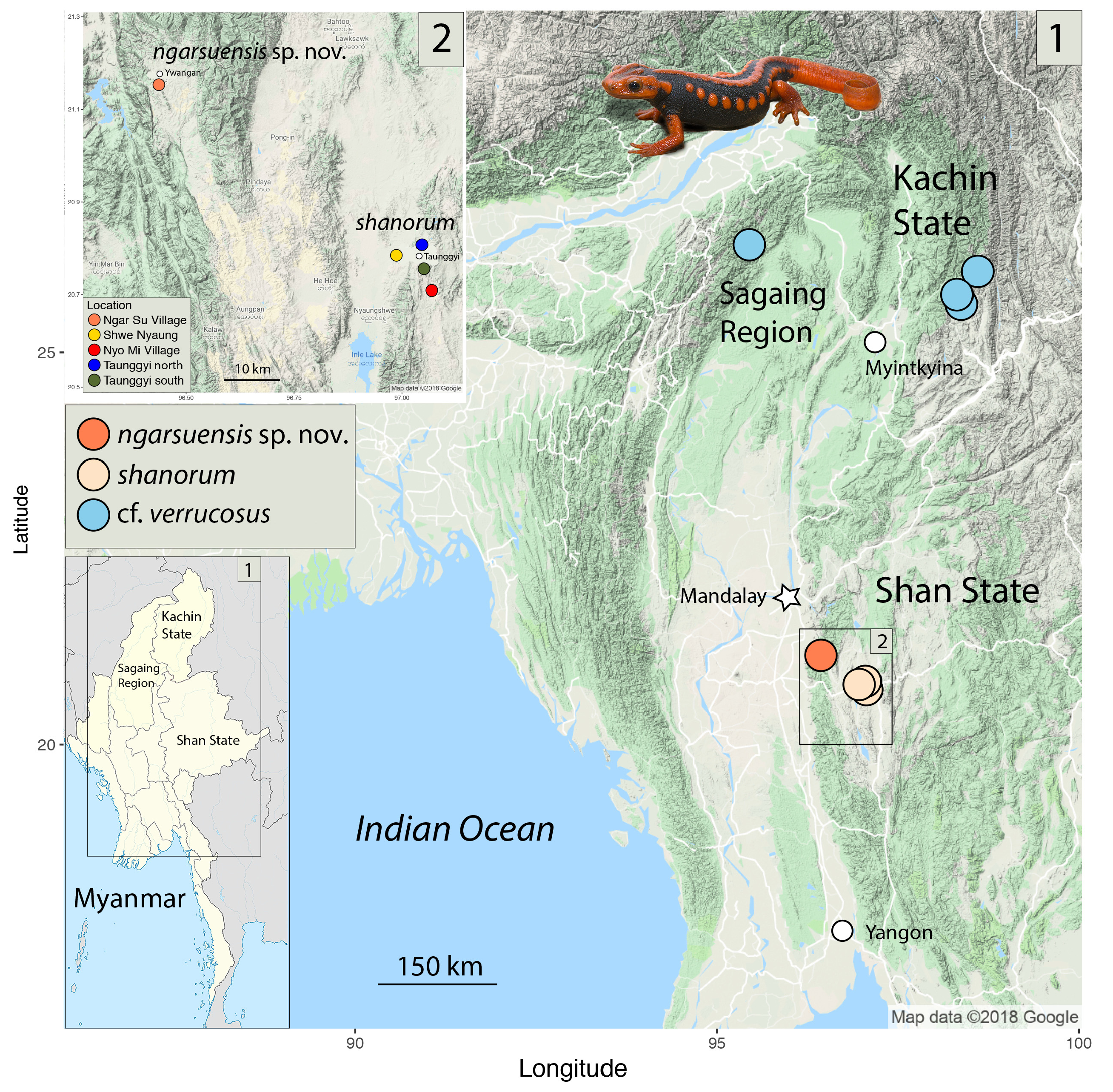
Holotype. Gravid female LSUHC 13762 View Materials collected on 24 October 2017 at 2330 hrs by Evan S. H. Quah, Robert E. Espinoza, Myint Kwaw Thura, and L. Lee Grismer from Baw Hto Chang in Ngar Su Village , Ywangan Township, Taunggyi District, Shan State, Myanmar (21.15364° N, 96.43660°E WGS84 ) at 1212 m in elevation. GoogleMaps
Paratypes. Adult male paratypes LUSHC 13763–64 bear the same collecting data as the holotype ( Table 4) GoogleMaps .
Additional referred material. A lot of 15 gilled larvae ( LSUHC 13761 View Materials ) ranging from 10–33 mm SVL (stage 44 of Grosse 2013) bearing the same locality data as the holotype were collected during the evening of 24 October 2017 and the morning of 25 October 2017 between 1000 and 1100 hrs .
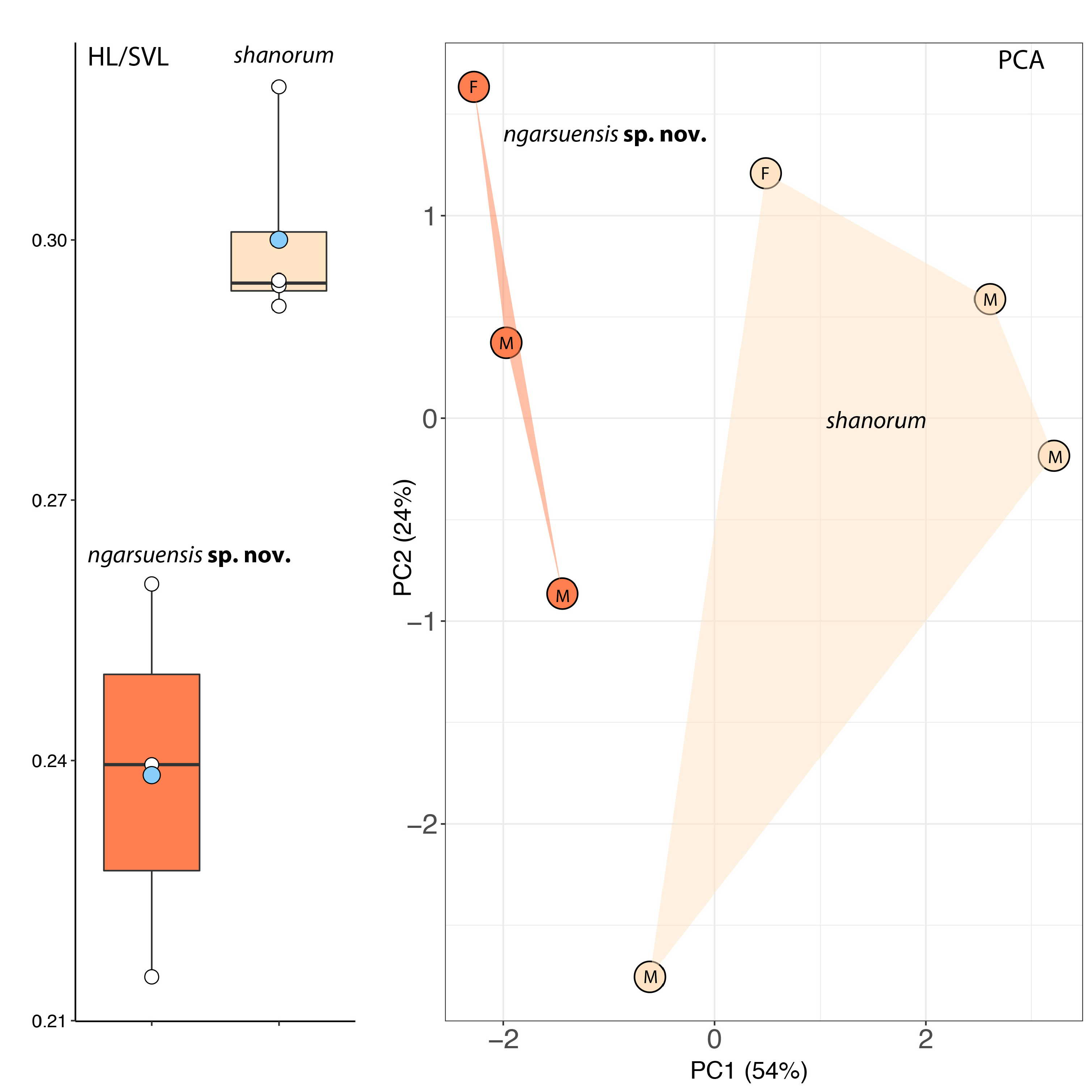
Diagnosis. A member of the genus Tylototriton based on its dark dorsal ground color with light-colored markings, well-developed cephalic and vertebral ridges, strong sexual dimorphism ( Dubois & Raffaëlli 2009), and its phylogenetic relationships ( Wang et al. 2018). A large-sized newt (female SVL = 102.3 mm; male SVL 74.9– 76.4 mm); head relatively short (HL/SVL = 0.22–0.26), subtriangular in dorsal profile; skin rough, glandular, bearing fine granules; dorsolateral boney ridges on head prominent; vertebral ridge low and wide; rib nodules relatively indistinct, sometimes lacking medially; vomeropalatine tooth series form an inverted, narrow V-shape, anteriorly projecting beyond the choanae but not in contact with them, teeth confined to palatine bone, not vomers; dorsal ground color black; anterior of head, paratoid region, vertebral ridge, rib nodules, limbs, and side of tail dark-brown, nearly black; labial regions, palms and soles, vent region, and subcaudal region light-brown.
Description of holotype. Habitus stout; head narrower than body, wider than long, subtriangularly shaped in dorsal profile, slightly sloping in lateral profile; snout short, rounded in dorsal profile, truncate in lateral profile, extending beyond lower jaw; nostrils on anterior margin of snout, facing forward, narrowly visible from above; labial fold absent; vomeropalatine tooth series form an inverted, narrow V-shape, anteriorly projecting beyond the choanae but not in contact with them, teeth confined to palatine bone, not vomers; tongue oval, attached to anterior floor of mouth but free posteriorly and laterally; no glandular ridge on midline of crown but short ridge on snout; dorsolateral bony ridges on head wide and slightly protruding, extending from lore to anterior end of parotoids, posterior ends turned slight toward midline; parotoids enlarged, crescent-shaped, projecting posteriorly with posterior margins curved medially; low, wide, weakly segmented, vertebral tubercular ridge extends from occiput onto anterior one-half of dorsal margin of tail, separated from ridge on midline of snout; dorsolateral row of weak glandular warts (rib nodules) on each side from level of axillae to level of posterior margin of vent (i.e. base of tail) on anterior margin of tail but not extending father down the tail; anterior and posterior nodules more prominent than the barely discernable medial series; approximately 15 rib nodules on both sides; skin rough; small, glandular warts on most dorsal and ventral surfaces; warts on crown, nape, and ventrolateral region in clusters of small glands, those on throat, tail, and ventral surfaces of limbs granular, those on belly arranged in transverse striations; pectoral region bearing a smooth, glandular, ovoid-shaped patch; gular fold prominent; adpressed limbs do not overlap; four fingers, five toes, all without webbing; and tail laterally compressed, tapering posteriorly, bearing sharp narrow dorsal fin most distinct posteriorly, a smooth ventral ridge, bluntly acuminate tip in lateral profile.
Color of holotype in life ( Fig. 4 View FIGURE 4 ). Dorsal ground color of top of head and trunk black; dorsal surfaces of limbs, and lateral surfaces of tail dark-brown; iris black; gular region, belly, and ventral surfaces of limbs dark-grey; anterior of head and parotoids dull-brown; rib nodules and vertebral ridge dark-brown, barely discernable from black trunk coloration; upper and lower lips, palms and soles, and subcaudal region light-brown.
Variation ( Figs 4 View FIGURE 4 , 5 View FIGURE 5 ). The male paratypes generally approximate the female holotype in coloration and pattern. LSUHC 13764 View Materials is darker overall than the holotype in that it is essentially black on all dorsal surfaces except for the upper lips, lores, and parotoids. The vertebral ridge is the same color as the trunk and the rib nodules are only weakly discernable by being slightly lighter in color. In LSUHC 13763 View Materials , these areas are only slightly lighter but they are more discernable than in the holotype or LSUHC 13764 View Materials . The middle series of rib nodules on the left side in LSUHC 13763 View Materials are barely discernable as in the holotype. The adpressed limbs greatly overlap in the paratypes but do not overlap in the holotype. The holotype measures 102.3 mm in SVL whereas the largest male ( LSUHC 13763 View Materials ) measures 76.4 mm. Other mensural differences are presented in Table 2 .
Distribution ( Fig. 1 View FIGURE 1 ). Tylototriton ngarsuensis sp. nov. is known only from Baw Hto Chang in Ngar Su Village, Ywnagan Township, Taunggyi District, Shan State, Myanmar (21.15364° N, 96.43660°E WGS84) although locals told us they can be found in other nearby streams and ponds.
Etymology. The specific epithet ngarsuensis is a toponym in reference to Ngar Su Village, the type locality.
Natural history. Baw Hto Chang is a shallow, slow-moving stream during the monsoon months (May- September) that runs through a narrow corridor of forest ( Fig. 6 View FIGURE 6 ). The area beyond the corridor has been converted to paddy fields and thus this narrow band of forest is likely to serve as the only place where newts can estivate during the colder and drier winter months. The adult newts were found in the late evening walking along the sandy bottom in clear water 1 m deep in the vicinity of underwater vegetation. Larvae were collected from shallower areas near the shore. We returned the next morning and found more larvae in the water along the edges of the shore, usually beneath leaves or other cover. The fact that we collected larvae as small as 10 mm SVL and a gravid female indicates newts are still breeding during late October. Most other species breed from April through July ( Bernardes et al. 2013, 2017) or from June through September in the case of Tylototriton himalayanus ( Dasgupta 1996) . Villagers from Ngar Su Village told us the newts are quite common on land during the months of May and June at the beginning of monsoon season at which time they are presumably migrating to the water. Villagers also say it is not uncommon for newts to wander into their houses during this period.
Comparisons. Tylototriton ngarsuensis sp. nov. is most closely related to T. shanorum but differs from it by having much larger adult females (102.3 mm vs 87.9 mm); a significantly shorter head in males (HL/SVL 0.22– 0.26 vs. 0.29–0.32; p = 0.041 for males; 0.22 vs 0.29 for females; Fig. 3 View FIGURE 3 ); having large, rounded rib nodules with diameters equivalent to or greater than that of eye as opposed to having small, slightly elongated rib nodules with diameters less than that of the eye; by having a thick, glandular, vertebral tubercular ridge as opposed to a narrow and less glandular ridge; 15 vs 14 rib nodules; paratoid ridge beginning posterior to the orbits as opposed beginning in the loreal region; top of head, vertebral ridge, rib nodules, limbs, and side of tail nearly black as opposed being red-brown; labial regions, palms, soles, and subcaudal region dark-brown as opposed to being dull-yellow; and venter and underside of limbs being dark-gray as opposed to dull-yellow. Tylototriton ngarsuensis sp. nov. and T. shanorum share an uncorrected pairwise sequence divergence between them of 3.0–3.4% which is commensurate with that among other well-established sister species ( Stuart et al. 2010).
Tylototriton ngarsuensis Color of holotype in life ( Fig. 4 View FIGURE 4 ). Dorsal ground color of top of head and trunk black; dorsal surfaces of limbs, and lateral surfaces of tail dark-brown; iris black; gular region, belly, and ventral surfaces of limbs dark-grey; anterior of head and parotoids dull-brown; rib nodules and vertebral ridge dark-brown, barely discernable from black trunk coloration; upper and lower lips, palms and soles, and subcaudal region light-brown.
Variation ( Figs 4 View FIGURE 4 , 5 View FIGURE 5 ). The male paratypes generally approximate the female holotype in coloration and pattern. LSUHC 13764 is darker overall than the holotype in that it is essentially black on all dorsal surfaces except for the upper lips, lores, and parotoids. The vertebral ridge is the same color as the trunk and the rib nodules are only weakly discernable by being slightly lighter in color. In LSUHC 13763, these areas are only slightly lighter but they are more discernable than in the holotype or LSUHC 13764. The middle series of rib nodules on the left side in LSUHC 13763 are barely discernable as in the holotype. The adpressed limbs greatly overlap in the paratypes but do not overlap in the holotype. The holotype measures 102.3 mm in SVL whereas the largest male (LSUHC 13763) measures 76.4 mm. Other mensural differences are presented in Table 2.
Distribution ( Fig. 1 View FIGURE 1 ). Tylototriton ngarsuensis sp. nov. is known only from Baw Hto Chang in Ngar Su Village, Ywnagan Township, Taunggyi District, Shan State, Myanmar (21.15364° N, 96.43660°E WGS84) although locals told us they can be found in other nearby streams and ponds.
Etymology. The specific epithet ngarsuensis is a toponym in reference to Ngar Su Village, the type locality.
Natural history. Baw Hto Chang is a shallow, slow-moving stream during the monsoon months (May- September) that runs through a narrow corridor of forest ( Fig. 6 View FIGURE 6 ). The area beyond the corridor has been converted to paddy fields and thus this narrow band of forest is likely to serve as the only place where newts can estivate during the colder and drier winter months. The adult newts were found in the late evening walking along the sandy bottom in clear water 1 m deep in the vicinity of underwater vegetation. Larvae were collected from shallower areas near the shore. We returned the next morning and found more larvae in the water along the edges of the shore, usually beneath leaves or other cover. The fact that we collected larvae as small as 10 mm SVL and a gravid female indicates newts are still breeding during late October. Most other species breed from April through July ( Bernardes et al. 2013, 2017) or from June through September in the case of Tylototriton himalayanus ( Dasgupta 1996) . Villagers from Ngar Su Village told us the newts are quite common on land during the months of May and June at the beginning of monsoon season at which time they are presumably migrating to the water. Villagers also say it is not uncommon for newts to wander into their houses during this period.
Comparisons. Tylototriton ngarsuensis sp. nov. is most closely related to T. shanorum but differs from it by having much larger adult females (102.3 mm vs 87.9 mm); a significantly shorter head in males (HL/SVL 0.22– 0.26 vs. 0.29–0.32; p = 0.041 for males; 0.22 vs 0.29 for females; Fig. 3 View FIGURE 3 ); having large, rounded rib nodules with diameters equivalent to or greater than that of eye as opposed to having small, slightly elongated rib nodules with diameters less than that of the eye; by having a thick, glandular, vertebral tubercular ridge as opposed to a narrow and less glandular ridge; 15 vs 14 rib nodules; paratoid ridge beginning posterior to the orbits as opposed beginning in the loreal region; top of head, vertebral ridge, rib nodules, limbs, and side of tail nearly black as opposed being red-brown; labial regions, palms, soles, and subcaudal region dark-brown as opposed to being dull-yellow; and venter and underside of limbs being dark-gray as opposed to dull-yellow. Tylototriton ngarsuensis sp. nov. and T. shanorum share an uncorrected pairwise sequence divergence between them of 3.0–3.4% which is commensurate with that among other well-established sister species ( Stuart et al. 2010).
Tylototriton ngarsuensis sp. nov. differs from all other species of Tylototriton as follows (data from Stuart et al. 2010; Hou et al., 2012; Nishikawa et al. 2013a,b, 2014; Khatiwada et al. 2015; Le et al. 2015; Phimmachak et al. 2015): from T. podicthys it differs by lacking orange coloration on the limbs and lateral surfaces of the tail and by lacking a boney ridge on the midline of the crown as opposed to having these traits; from T. anguliceps and T. shanjing it differs by lacking a boney ridge on the midline of the crown as opposed to the presence of a boney ridge; from T. anguliceps it differs by lacking an orange head, limbs, tail, parotoids, rib nodules, and vertebral ridge as opposed to having these structures being orange; from T. uyenoi and T. pulcherrima it differs by lacking orangecolored limbs, tail, parotoids, rib nodules, and vertebral ridge as opposed to having these structures colored orange; from T. kweichowensis and T. pseudoverrucosus it differs by having isolated rib nodules as opposed to the rib nodules being connected and forming dorsolateral ridges; from T. shanjing it differs by having no sharp contrast between the orange crown of the head and black nape as opposed to a sharp contrast, having dark coloration on ventral surfaces of limbs versus orange coloration, and the tips of the digits being dark as opposed uniformly orange; from T. taliangensis it differs by having as opposed to lacking rib nodules; from T. verrucosus it differs by lacking a glandular ridge on the midline of the crown as opposed to having a ridge on the crown; from T. yangi it differs by having uniformly orange parotoids as opposed to the light-colored parotoid regions being restricted to the posterior margins of the glands; and from T. himalayanus it differs in having the head longer than wide, a truncate snout in lateral profile as opposed to a rounded snout, and ventral surfaces being dark-gray as opposed to cream-colored. differs from all other species of Tylototriton as follows (data from Stuart et al. 2010; Hou et al., 2012; Nishikawa et al. 2013a,b, 2014; Khatiwada et al. 2015; Le et al. 2015; Phimmachak et al. 2015): from T. podicthys it differs by lacking orange coloration on the limbs and lateral surfaces of the tail and by lacking a boney ridge on the midline of the crown as opposed to having these traits; from T. anguliceps and T. shanjing it differs by lacking a boney ridge on the midline of the crown as opposed to the presence of a boney ridge; from T. anguliceps it differs by lacking an orange head, limbs, tail, parotoids, rib nodules, and vertebral ridge as opposed to having these structures being orange; from T. uyenoi and T. pulcherrima it differs by lacking orange-colored limbs, tail, parotoids, rib nodules, and vertebral ridge as opposed to having these structures colored orange; from T. kweichowensis and T. pseudoverrucosus it differs by having isolated rib nodules as opposed to the rib nodules being connected and forming dorsolateral ridges; from T. shanjing it differs by having no sharp contrast between the orange crown of the head and black nape as opposed to a sharp contrast, having dark coloration on ventral surfaces of limbs versus orange coloration, and the tips of the digits being dark as opposed uniformly orange; from T. taliangensis it differs by having as opposed to lacking rib nodules; from T. verrucosus it differs by lacking a glandular ridge on the midline of the crown as opposed to having a ridge on the crown; from T. yangi it differs by having uniformly orange parotoids as opposed to the light-colored parotoid regions being restricted to the posterior margins of the glands; and from T. himalayanus it differs in having the head longer than wide, a truncate snout in lateral profile as opposed to a rounded snout, and ventral surfaces being dark-gray as opposed to cream-colored.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |