
Aeolosaurus maximus, Santucci, Rodrigo M. & De, Antonio C., 2011
|
publication ID |
https://doi.org/10.5281/zenodo.205522 |
|
DOI |
https://doi.org/10.5281/zenodo.5625343 |
|
persistent identifier |
https://treatment.plazi.org/id/03AE87A9-FF8E-FF87-FF4C-FD8AE3F0FABD |
|
treatment provided by |
Plazi |
|
scientific name |
Aeolosaurus maximus |
| status |
sp. nov. |
Aeolosaurus maximus sp. nov.
1999a Aeolosaurus sp. Bertini et al.
1999b Aeolosaurus sp. Bertini et al.
2001 Aeolosaurus sp. Santucci and Bertini, p. 308, Fig. 2 View FIGURE 2 A
Derivation of name. maximus , in reference to the size of the specimen, meaning large in Latin.
Holotype. MPMA 12-0001-97, two incomplete posterior cervical vertebrae; seven incomplete cervical ribs; a fragmentary anterior dorsal centrum; a probable fragment of a middle dorsal vertebra; a fragmentary posterior dorsal vertebrae; several incomplete diapophysis of dorsal vertebrae; 12 incomplete dorsal ribs; six articulated anterior caudal vertebrae; a mid caudal centrum; two posterior caudal vertebrae; six anterior, one mid, and one posterior haemal arches; a probable fragmentary scapula; an incomplete right humerus; a probably fragmentary left humerus; a probably incomplete radius; incomplete right femur; left femur; left ischium; and several unidentified fragments. The material is housed in the Museu de Paleontologia de Monte Alto (Monte Alto Paleontological Museum), São Paulo, Brazil.
Locality and horizon. Reddish, massive sandstones locally with carbonatic cementation, top of the Adamantina Formation, Campanian–Maastrichtian, Upper Cretaceous, approximately 12 Km SW of the city of Monte Alto, São Paulo State, Brazil ( Fig. 1 View FIGURE 1 ).
Diagnosis. Titanosaur characterized by the following unique association of characteristics: well-developed posterior protuberance below the articular area on the anterior and middle haemal arches and lateral bulge on the distal portion of the articular process of the mid-posterior haemal arches. An additional set of ambiguous synapomorphies includes: presence of posterior centrodiapophyseal lamina (pcdl) at least 50% thicker than the postzygodiapophyseal lamina (podl) in posterior cervical vertebrae; posterior dorsal vertebrae with intrapostzygapophyseal lamina and an oblique anterior centropostzygapophyseal lamina (acpol) which bifurcates from the proximal portion of the centropostzygapophyseal lamina (acpol); mid-thoracic ribs bearing well-developed anterior and posterior crests with a D-shaped cross section. These features are considered ambiguous because they cannot be assessed in other Aeolosaurus species so far, and may correspond to a set of synapomorphies of Aeolosaurus or even Aeolosaurini.
Taphonomic note. The bones of A. maximus were unearthed from a small area of about 100m 2 ( Fig. 1 View FIGURE 1 ) and from the same stratigraphical level. No repeated elements were found. According to their position in the field, they represent a single individual that was lying with its left side down. The two fragmentary posterior cervical vertebrae together with several pairs of cervical ribs were found articulated, with the cervical ribs overlapping at least two consecutive ones. Their position in the field suggests the neck was dorsiflexed, which indicates the skeleton has undergone at least a slightly post-mortem exposure prior its final burial. The same pattern was observed in the recovered anterior caudal series. The high degree of articulation, together with the presence of both large complete bones and small and delicate elements, also suggests the skeleton has undergone little transport.
Several theropod and crocodylomorph teeth have been recovered from the area where both femora were found. The presence of these teeth around this portion of the skeleton suggests that small theropod dinosaurs and crocodylomorphs have fed on the carcass. This pattern has also been reported elsewhere, where teeth of carnivorous dinosaurs are found near the sacral region of articulated or semi-articulated sauropods ( Buffetaut and Suteethorn 1989). However, bite marks have not been found in the bones.
Description. Cervical vertebrae. Comparing the cervical vertebrae of Aeolosaurus maximus with the nearly complete cervical series of Malawisaurus dixeyi ( Gomani 2005) and the Series A from Peirópolis ( Powell 2003), they seem to correspond to posterior cervical vertebrae, probably the 10th and 11th in the series. The two articulated posterior cervical vertebrae consist of the left lateral portion of the neural arches lacking the centra and the neural spines. Because of their large size (the preserved portion of the posteriormost cervical vertebra is 85 cm in length) and their slender structure, these vertebrae were kept in articulation during preparation. Moreover, they also have signs of dorsoventral deformation ( Fig. 2 View FIGURE 2 ).
According to the preserved portions, the internal bony tissue (camellae) of the centra is composed of subcentimetrical coels circumscribed by thin lamellae. Both vertebrae have the left cervical ribs attached to their centra. The capitulum and tuberculum are thin sheets of bone which are anteroposteriorly wide. However, their contact with the transverse processes is not clear. The tuberculum is internally reinforced by a columnar ridge that extends from the base of the capitulum until the tuberculum. The anterior end of the cervical ribs is shorter than the posterior one and tapers to a point. The posterior end is long and slender, formed by a thin dorsoventrally convex sheet of bone that gradually becomes oval in cross section toward the end. According to the preserved cervical ribs (some of them are 60 cm in length) and their position in the field, they may overlap two or three adjacent cervical vertebrae ( Fig. 1 View FIGURE 1 ).
On both vertebrae the parapophyses are badly damaged and therefore cannot be properly described. The diapophyses are blade-like projections and lay under the level of the prezygapophyses. On the posterior margin of the diapophyses there is a small sheet of bone that projects posteriorly. The most striking feature of these vertebrae is the strong development of the posterior centrodiapophyseal lamina, which is stouter than the postzygodiapophsyeal lamina. Although not completely preserved on both centra, the posterior centrodiapophyseal lamina seems to extend until the posterior margin of the centrum. The postzygodiapophsyeal lamina is thin, well-developed, and extends until the anterior margin of the postzygapophysis. This region is only preserved in the anteriormost cervical vertebra, which is broken and levered forward and upward by the forward displacement of the proceeding cervical vertebra ( Fig. 2 View FIGURE 2 ).
The prezygapophysis is short and robust, with a wide, flat and elliptical articular facet. The anterior portion of the neural spine (left spinoprezygapophyseal lamina) is preserved. It originates right at the posterior margin of the prezygapophyseal articular facet and extends upward and backward ( Fig. 2 View FIGURE 2 ).
Dorsal vertebrae. Only fragmentary dorsal vertebrae are preserved. A fragmentary neural arch is preserved, comprising a partial neural spine, the right diapophysis, and the right prezygapophysis. According to the height of the neural spine and its shape, this dorsal vertebra is regarded as a middle dorsal vertebra. The neural spine is tall and has an acute triangular shape in anterior view and is anteroporsteriorly short. The diapophysis is robust, short, and directed laterally. The spinodiapophyseal lamina is stout and well developed. The prezygapophysis is broken and seems to be shifted to a more medial position because its articular facet is unusually aligned to the sagittal plane ( Fig. 3 View FIGURE 3 A). A dorsal centrum ( Fig. 3 View FIGURE 3 B1-2) is poorly preserved. Its shape cannot be established, but it has welldeveloped internal coels. In the anterior face of the neural arch there are three fossae that widen into large internal chambers. Between these fossae, on the anterior portion of the neural arch, there are two diagonal shallow ridges that cross each other near their base. Some isolated transverse processes also have spongy bony tissue.
A left postzygapophysis is attributed to an anterior dorsal vertebra. It is robust and has a wide and flat articular facet. On its medial side a thin lamina is attached to it, which seems to be a fragmentary intrapostzygapophyseal lamina.
A large fragmentary posterior dorsal vertebra is preserved and consists of part of the neural arch fused to the centrum, both postzygapophyses, and part of the postzygodiapophsyeal lamina. The dorsal margin of the posterior articulation of the centrum seems to form a large concavity, which indicates that the centrum was opisthocoelous.
The neural canal is large and semi-oval in shape. In lateral view the posterior portion of the neural arch seems to bifurcate distally, where the side that forms the lateroposterior portion of the neural arch (cpol) bifurcates from the base of the postzygapophysis downward. Because of that, an additional oblique lamina, here called anterior centropostzygapophyseal lamina (acpol), originates from the base of the postzygapophysis and extends forward ( Fig. 3 View FIGURE 3 C1-2). Unfortunately, its proximal end is not preserved and, therefore, the area where it attaches is not known. The postzygodiapophsyeal lamina is thin and well-developed. The postzygapophyses are robust with large and flat articular facets and face ventrally. They are supported by simple, robust, and well-developed spinopostzygapophyseal laminae. The postspinal lamina consists of a shallow and stout ridge. Although the hyposphene is absent, the postzygapophyses are linked to each other by a thin intrapostzygapophyseal lamina at their bases ( Fig. 3 View FIGURE 3 C1-2).
Caudal vertebrae. The anterior caudal vertebrae consist of a series of six articulated elements. By comparison with complete caudal series from Brazil (Kellner et al. 2005) and other anterior caudal vertebrae of other Aeolosaurus specimens ( Salgado et al. 1997a), they would correspond to the fourth to ninth caudal vertebra ( Table 1 View TABLE 1 ). All caudal vertebrae are strongly procoelous, with the apex of the convexity of the posterior end slightly displaced above the midline of the centrum, and lack internal spongy bony tissue ( Figs. 4–5 View FIGURE 4 View FIGURE 5 ).
Distance between the articular facets of the prezyg./postzyg. 21,5 20,7 - - 17,5 18,0 Width at the tip of transverse processes - 34,0* 30,4* 30,0* - - The anteriormost preserved caudal vertebra (probably the fourth caudal vertebra) lacks the right transverse process, the right prezygapophysis, and the distal end of the neural spine. The centrum is relatively long and higher than wide. The ventral face is slightly concave and surrounded by shallow lateral ridges. The neural arch is anteroposteriorly short and located on the anterior half of the centrum. The transverse process is stout and directed backward. A large protuberance is located between the anterior margin of the base of the transverse process and the prezygapophysis ( Fig. 4 View FIGURE 4 B). The prezygapophysis is robust, directed forward, and slightly curved downward when seen in lateral view. The postzygapophyses are large, located at the base of the neural arch, and have elliptical articular facets. At least the base of the neural spine is compressed laterally. The well-developed spinoprezygapophyseal laminae are stout and extend over the preserved portion of the neural spine ( Fig. 4 View FIGURE 4 A–B).
The fifth caudal vertebra is nearly complete only lacking the distal end of the neural spine. The centrum is higher than wide and the lateral faces are concave anteroposteriorly. The posterior articulation of the centrum has a shallow elliptical groove. The morphology of this caudal vertebra is similar to that of the previous one, except for the curvature of the prezygapophysis, which is less developed, and the distal end of the transverse process, which is slightly expanded dorsoventrally ( Fig. 4 View FIGURE 4 C–D).
The four remaining anterior caudal vertebrae are well preserved except for the last two vertebrae which lack the transverse processes. The centra are higher than wide and become progressively longer toward the posterior caudal vertebrae. In posterior view, the posterior ends are roughly hexagonal in shape. The articular area for the haemal arches is well-developed, forming a protuberance with a shallow pit at the ventroposterior margin of the centra. The ventral faces are slightly concave anteroposteriorly and laterally compressed, with incipient lateroventral ridges. The transverse processes are robust and strongly directed backward. The neural arches are located on the anterior half of the centra, reaching the anterior margin of the centra of the last two preserved anterior caudal vertebrae. The prezygapophyses become straight and relatively long, with moderately developed articular facets. The spinoprezygapophyseal laminae are well-developed, forming a deep fossa at the base of the neural spine. The postzygapophyses are robust, with concave articular facets and stout spinopostzygapophyseal laminae. The neural spines are laterally compressed at their base and laterally expanded at their distal end. They are directed forward, mainly in the last two anterior caudal vertebrae. The prespinal and postspinal laminae are incipiently developed ( Fig. 4 View FIGURE 4 E–J).
The partial mid-caudal centrum is poorly preserved and, therefore, cannot provide any further information.
The two posterior caudal vertebrae are well preserved. The centra are procoelous and wider than high. The ventral and lateral faces are gently concave anteroposteriorly. The neural arches are anteroposteriorly short and located on the anterior half of the centra. The transverse processes are absent. The prezygapophyses have rudimentary articular facets and are directed forward. The neural spines are low, anteroposteriorly long, and transversally compressed. The postzygapophyses are poorly developed, consisting of shallow articular facets on the ventral margin of the neural spine ( Fig. 4 View FIGURE 4 K).
Dorsal ribs. Several incomplete dorsal ribs have been recovered. Unfortunately, they were not found in their anatomical position and, therefore, their assignment as anterior or posterior ribs have been made on the basis of better preserved sauropod material such as Apatosaurus , Camarasaurus , Brachiosaurus , and Opisthocoelicaudia . The more robust and platelike fragments were regarded as the anterior ribs, whereas the slender ribs were considered as the posterior ones ( Fig. 6 View FIGURE 6 B–E). None of the anterior ribs have the capitulum and tuberculum well preserved ( Fig. 6 View FIGURE 6 B–C). In both anterior and posterior dorsal ribs, the proximal portion consists of spongy bony tissue, mainly in a large rib fragment, where the coels open externally forming an elliptical pit. A fragment of proximal end is considered to be the first or second dorsal rib ( Fig. 6 View FIGURE 6 C). It is a blade-like element with the proximal end directed anteriorly. The posterior portion is damaged and exposes the internal spongy bony tissue and, because of that, it is not possible to determine the shape of the cross section. Other fragments regarded as anterior dorsal ribs have the cross section of the proximal end triangular in shape. The mid-dorsal ribs have shafts with subtriangular cross sections that become blade-like or elliptical toward their distal ends. A mid-thoracic rib has a marked depression between the capitulum and the tuberculum in medial view. The proximal portion of the shaft is D-shaped in cross section with sharp well-developed ridges on both the antero and posterolateral margins ( Fig. 6 View FIGURE 6 D). The best-preserved rib is about 900 mm in length and is considered to be a posterior one. It only lacks part of the capitulum and the tuberculum and, probably, part of the distal end. It is straight in medial view and gently curved in posterior view. It has the proximal portion of the shaft flat and is anteroposterioly concave in medial view. However, the distal end becomes triangular in cross section as in the middle posterior ones ( Fig. 6 View FIGURE 6 E).
Haemal arches. Six anterior haemal arches are preserved ( Fig. 7 View FIGURE 7 A–F). Although not found articulated with the six anterior caudal vertebrae, some of them fit exactly in these caudal vertebrae. They are ‘Y’-shaped and open proximally, in the proximal chevrons the haemal canal is less than 50% of the length of the whole bone ( Table 2). Both proximal and distal processes are laterally compressed (but not laminar) mainly on the distal portion of the distal processes. In lateral view they gently curve backward. The top of the proximal process is nearly flat and has a large concave area in the posterodorsal margin, forming double articular facets. In lateral view this depression is ventrally delimited by a posterior protuberance. The distal process has a rounded distal end in lateral view and is anteroposteriorly expanded. In posterior view the distal process has a thin ridge that runs through its whole posterior aspect ( Fig. 7 View FIGURE 7 A–F).
A haemal arch from a mid-caudal vertebra is well preserved ( Fig. 7 View FIGURE 7 G). It is ‘Y’-shaped with the haemal canal more than 50% of the length of the whole bone. Only the distal end of the distal process is laterally compressed. In lateral view it is nearly straight with the distal end curved backward. As the anterior haemal arches, the articulation with the centrum has a large posterior concave area which, in lateral view, is ventrally marked by a well-developed protuberance. The proximal process has a lateral protuberance located on its distal half ( Fig. 7 View FIGURE 7 G).
The distal haemal arch is a ‘V’-shaped element. The proximal processes have elliptical cross section with a mildly-developed protuberance on their posterolateral margins. The proximal articulations with the centrum have a flat top and a posterior concavity, forming the double articular facets. However, the posterior protuberance that delimitates its ventral margin is less developed than in the previous haemal arches ( Fig. 7 View FIGURE 7 H).
ha1 ha2 ha3 ha4 ha5 ha6 ha7 ha8 Total height 36,5* 38,2 - - - - 16,5 11,3 Height of haemal canal 18,7 17,7 17,2 16,5 - - 9,1 7,4 Proximal width 12,5 11,8 10,1 11,4 - - 9,2 8,1 Scapula. A large platelike fragment is considered to be part of the distal end of a scapula. It has a rounded dorsal margin and its thickness decreases toward the ventral margin.
Humeri. Both humeri are preserved ( Fig 8 View FIGURE 8 A–B). The right humerus (preserved length of 89 cm) lacks the distal end and the laterodorsal corner ( Fig. 8 View FIGURE 8 A). The proximal end has a well-developed anterior concavity and is medially expanded. The left one (preserved length of 63 cm) lacks both epiphyses, is anteroposterioly compressed, and is also badly damaged on its anterior portion ( Fig 8 View FIGURE 8 B).
Radius. A long bone element (preserved length of 56 cm) found near the humerus is interpreted as a radius. Unfortunately, it is badly damaged and strongly compressed, so that nothing can be said about its morphology ( Fig 8 View FIGURE 8 C).
Ischium. The left ischium is partially preserved ( Figs. 9 View FIGURE 9 B1–2, 10). It is a platelike element with a concave dorsoposterior margin. Both iliac and pubic pedicles are not completely preserved. However, the pubic articulation seems to be well-developed. The acetabular area is partially preserved, corresponding to a gently concave area between the pubic and iliac articulations, where the bone is thinner than the posterodorsal margin. According to the preserved portion of the distal process, it seems to be relatively long and its thickness decreases from the dorsal margin to the ventral one. The distal process is twisted medially which indicates that when in articulation with its counterpart they should have met in a horizontal plane ( Figs. 9 View FIGURE 9 B1–2).
Femur. Both femora were recovered ( Fig. 9 View FIGURE 9 A1–4), the left one is complete and well preserved, the right one lacks the distal end and part of the shaft. Although large (1.55 m in length), the femur is a relatively slender element. It is straight and strongly compressed anteroposteriorly, so that the shaft has an elliptical shape in cross section. The femoral head is well developed and extends well above the level of the greater trochanter and medially. The femoral head is not aligned with the great trochanter, so that in dorsal view it is slightly directed backward, forming a shallow concave area between both the great trochanter and the femoral head. A lateral bulge is present right under the great trochanter; it is gently convex and extends over the third proximal portion of the shaft. The fourth trochanter is relatively well-developed and located on the posteromedial portion of the shaft at its third upper part. The distal articulation is well-developed and extends from the posterior to the anterior margin of the femur so that in both posterior and anterior view there is a concave area between the tibal and fibular condyles. The same holds true for the distal margin. The tibial condyle is more developed than the fibular one in the way the posterior portion of the former is more developed posteriorly. However, the fibular condyle is stouter and bifurcates into two condyles. The internal (medial) one is more developed and slightly directed laterally. The lateral fibular condyle is less developed and forms part of the lateral corner of the distal end of the femur ( Fig. 9 View FIGURE 9 A1–4).
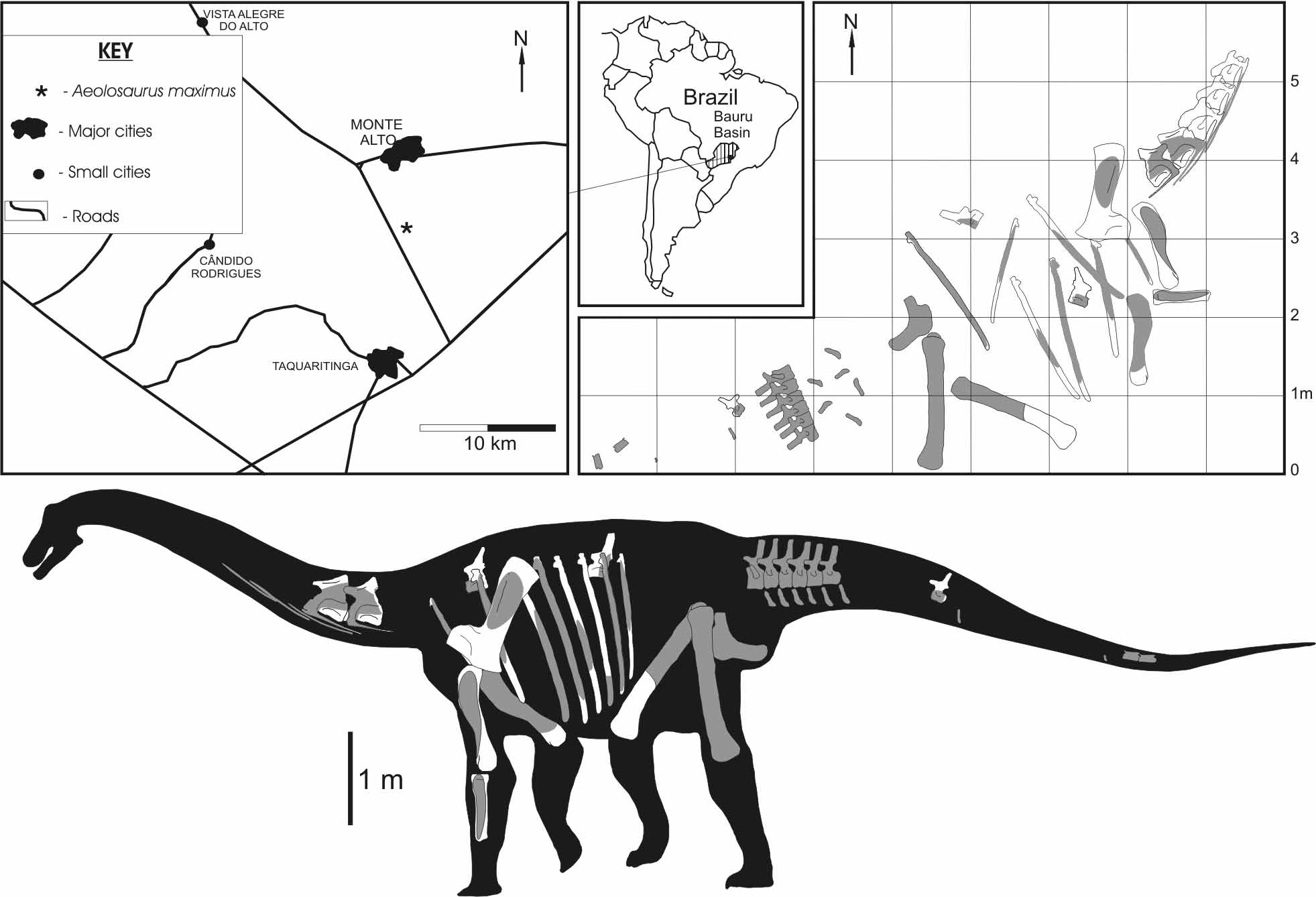
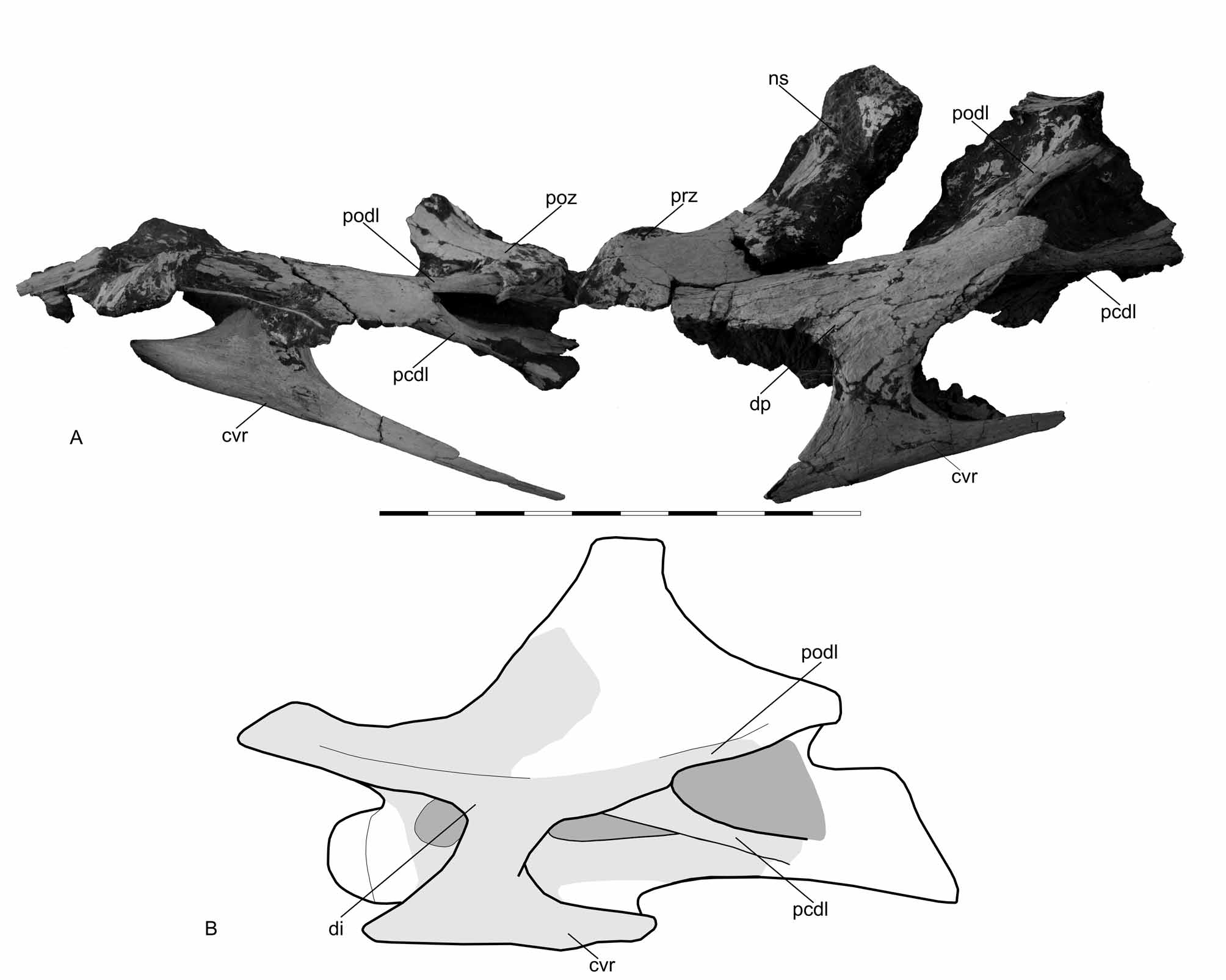


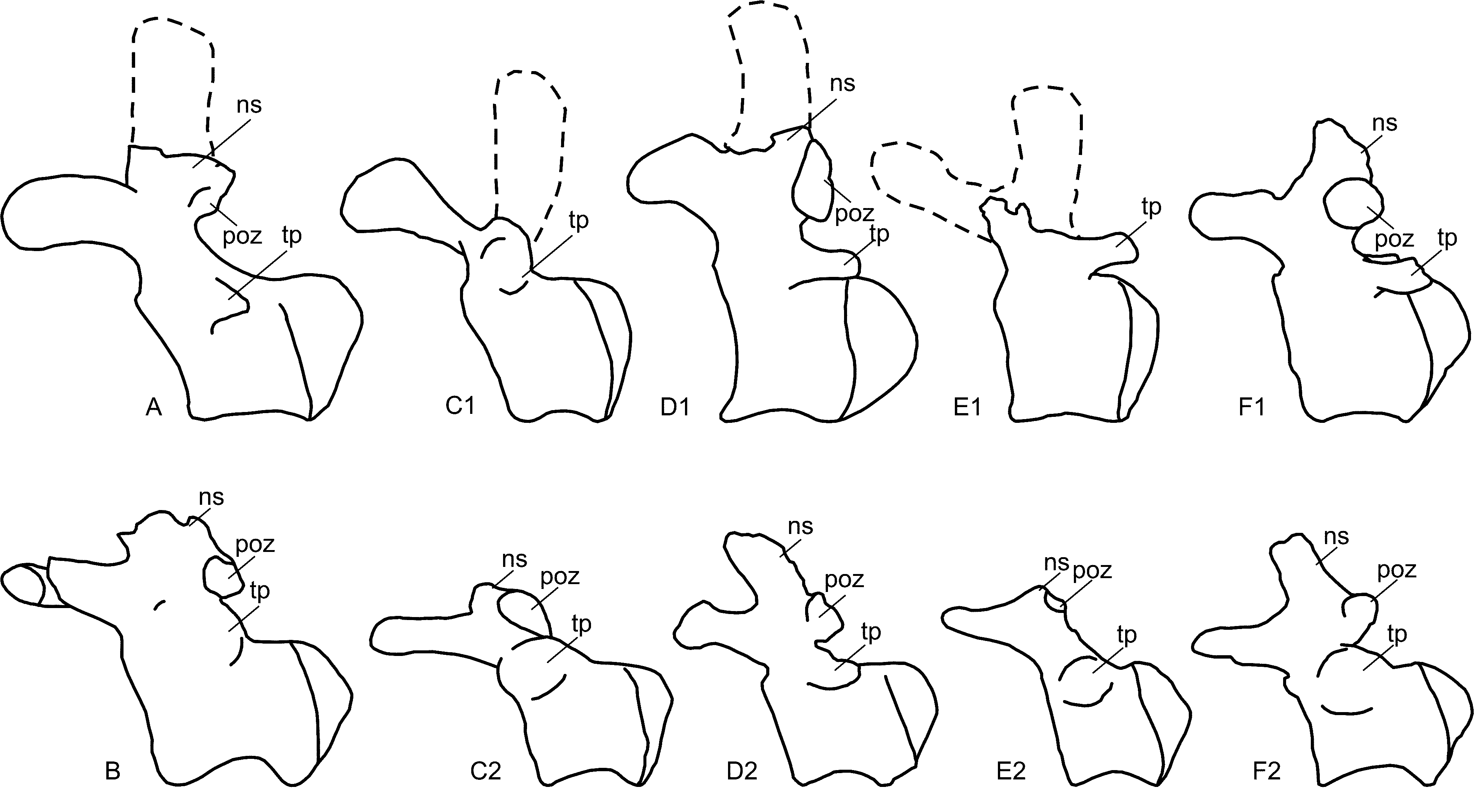



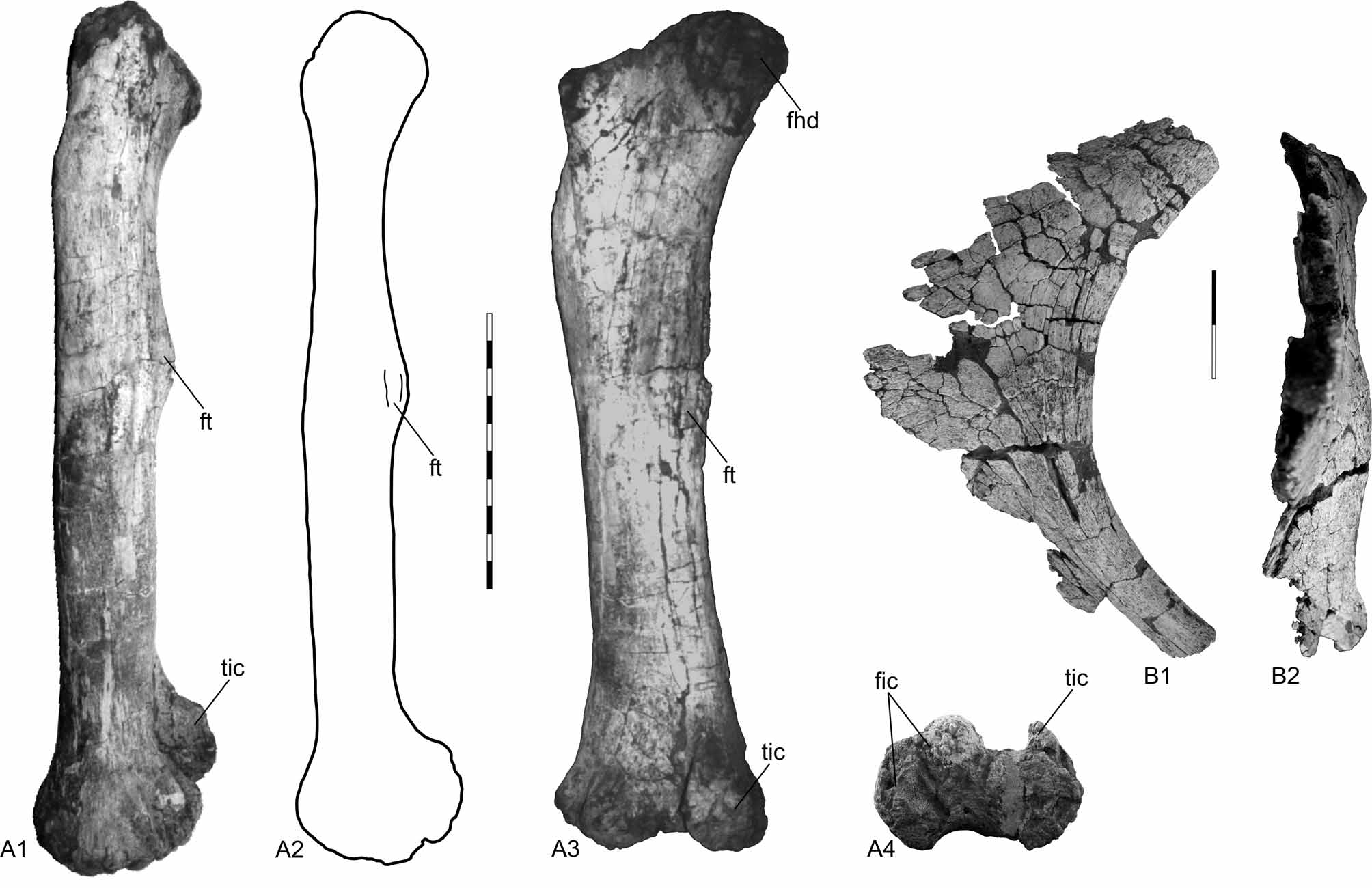
TABLE 1. Measurements of the preserved anterior caudal vertebrae of A. maximus (MPMA 12 - 0001 - 97). Measurements are in cm. Asteriscs indicate estimated measurements.
| C4 | C5 | C6 | C7 | 86 | C9 | |
|---|---|---|---|---|---|---|
| Total length including prezygapophises | 34,2 | 35,8 | 32,2 | 33,5 | 33,0 | 33,5 |
| Length of centrum (with condyle) | 26,3 | 25,5 | 24,0 | 20,4 | 22,0 | 22,5 |
| Length of centrum (without condyle) | 18,5 | 17,0 | 17,5 | 16,0 | 15,5 | 16,5 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |