
Hyloconis Kumata, 1963
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3594.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:B00799F3-F397-438C-B1E1-A8440E636921 |
|
DOI |
https://doi.org/10.5281/zenodo.6308334 |
|
persistent identifier |
https://treatment.plazi.org/id/03ADE350-B104-FF93-F1CF-FAAF8B0FCF39 |
|
treatment provided by |
Felipe |
|
scientific name |
Hyloconis Kumata, 1963 |
| status |
|
Hyloconis Kumata, 1963 View in CoL
Hyloconis Kumata 1963: 28 View in CoL .
Type species: Hyloconis puerariae Kumata, 1963 View in CoL , by original designation.
Historic account. Hyloconis includes four Japanese species: H. pueraria Kumata, 1963 , H. desmodii Kumata 1963 , H. lespedezae Kumata, 1963 , and H. wisteriae Kumata, 1963 , all off which feed on Fabaceae . Kumata (1963) proposed the genus because of differences in the biology and wing venation from those of Cremastobombycia , Neolithocolletis , and Phyllonorycter . The male genitalia of Hyloconis possess an appendage at the base of the valval costa and an incomplete transtilla ( Kumata 1963; Kuznetzov & Baryshnikova 2001 ). Kumata (1993) drew attention to the diagnostic value of the apical setae on the tegumen. Hyloconis has more than four setae on the apex of tegumen, similar to Cremastobombycia and different from Phyllonorycter (apex of tegumen without setae) and Neolithocolletis (four setae) ( Kumata 1993). The fifth species presently belonging to Hyloconis , H. improvisella ( Ermolaev, 1986) , was described from the Far East of Russia in the genus Lithocolletis . In the same paper, Ermolaev (1986) transferred the type species of Hyloconis , H. puerariae , to Lithocolletis , thus synonymizing Hyloconis under Lithocolletis . However, Park (1983) and Kamijo (1990) considered Hyloconis a valid genus. Later, Noreika (1994) placed L. improvisella to Hyloconis , and returned to Hyloconis two other species, H. puerariae and H. lespedezae , originally described by Kumata (1963) in Hyloconis . In keys and catalogues published later, Hyloconis was listed as one of the genera of Palaearctic Lithocolletinae ( Noreika 1994, 1997; Kuznetzov & Baryshnikova 1998 , 2001; Baryshnikova 2002 ; De Prins & De Prins 2005, 2012).
Until now, Hyloconis was known exclusively from the Palaearctic. Here we report Hyloconis luki De Prins , n. sp., from Central Africa.
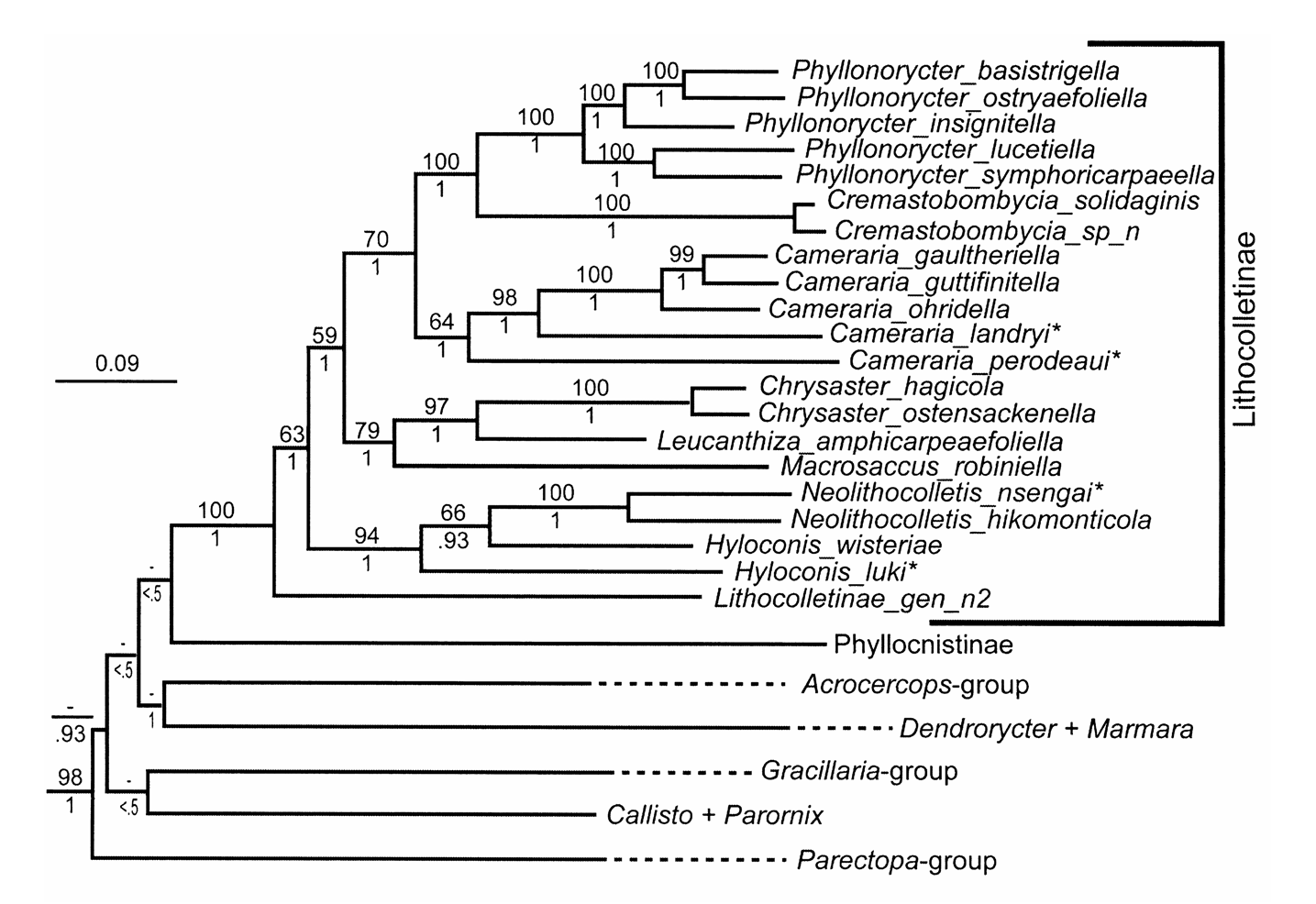
Diagnosis. According to Kumata (1963), Hyloconis is morphologically most closely related to Cremastobombycia and Phyllonorycter , but it also shares many external and internal characters with other Lithocolletinae genera. The adult head has a tuft of long scales that projects forward above the vertex between the antennae as in Cameraria , Neolithocolletis , Phyllonorycter , and Porphyrosela ; Chrysaster , Leucanthiza , and Protolithocolletis all possess a smooth vertex. The black margins of the white forewing markings in Hyloconis clearly differ from of comparable markings in Cameraria and Cremastobombycia . In the latter two genera the black margins of white forewings markings are situated apically whereas in Hyloconis they can be situated both basally and apically as in Chrysaster , Phyllonorycter , Porphyrosela , and Protolithocolletis . Forewing venation is similar to that of Cremastobombycia , Leucanthiza , Neolithocolletis , and Protolithocolletis , having veins R 2 and M 2 absent. Hindwing venation is similar to that of Cremastobombycia , Porphyrosela and Protolithocolletis in having M 2. The male genitalia are symmetrical or asymmetrical ( Kumata 1963: 28). In the male genitalia of Hyloconis , the apex of the tegumen has 4 or more setae, as in Neolithocolletis and Cremastobombycia , but differers from Phyllonorycter , which lacks setae on the apex of the tegumen. Other Lithocolletinae genera, such as Cameraria , Chrysaster , Macrosaccus and Porphyrosela , have one pair of setae on the apex of the tegument. The transtilla in Hyloconis is incomplete as in most species of Cameraria ; the valva has an ovate knob at the base of of the costa; the saccus is well developed, projected; and the anellus is sclerotized as in some Cameraria and Cremastobombycia . Sternum VIII in the male is enlarged into a wide flap. The female genitalia have a sclerotized plate on segment VIII, except in H. wisteriae Kumata, 1963 ; the ostium bursae opens in segment VIII, and the sterigma around the ostium bursae is sclerotized. The antrum and/or ductus bursae is usually sclerotized posteriorly. The corpus bursae is in the form of an enlarged, irregularly shaped sac, without signa or with a single very long signum (in H. wisteriae ). Larvae of Hyloconis make entirely flattened mines, different from the tentiform mines of Phyllonorycter and other lithocolletine genera. The cocoon of Hyloconis is orbicular, with frass placed on one side of mine in a round patch ( Kumata 1963). Hyloconis species feed on Fabaceae , a character shared with Chrysaster , Neolithocolletis , Porphyrosela and Protolithocolletis , and different from Cremastobombycia which feed on Asteraceae and Verbenaceae . The chaetotaxy of the last instar larva of Hyloconis is very similar to that of Neolithocolletis , but the thoracic legs of the last instar are not reduced in Hyloconis ( Kumata 1993) . Whereas Hyloconis may be paraphyletic with respect to Neolithocolletis ( Fig. 4 View FIGURE 4 ), support for H. wisteriae + Neolithocolletis in our analyses was weak (BP = 66%; PP = 0.93), and thus we consider the paraphyly of Hyloconis uncertain. Additional taxa will need to be sequenced to test the monophyly of Hyloconis .
Diagnosis of Afrotropical Hyloconis . Hyloconis is represented by only one species in the Afrotropical region, H. luki De Prins , n. sp., which is very small species with a wing length of ca. 1.5 mm. The external characters of Afrotropical Hyloconis are similar to those of Japanese Hyloconis species (2 paratypes of H. wisteriae and 2 paratypes of H. puerariae were compared, drawer Mi 10022 in BMNH, see also Kumata 1963: pl. 5, Figs 66–69 View FIGURES 63–68 View FIGURES 69–74 ). However, Afrotropical Hyloconis has strikingly different male genitalia from the Palaearctic species. Genitalia dissections are necessary for distinguishing nearly all Afrotropical lithocolletine genera.
Head: With tufted scales on posterior part of vertex and occiput; frons smooth, white, with silvery gloss. Maxillary palpus rudimentary; labial palpus short, drooping, filiform, pointed; haustellum of moderate length. Antenna nearly as long as forewing, flagellum thicker than in Phyllonorycter ; pedicel slightly thicker, but shorter than flagellomere; scape short and thick, with pecten.
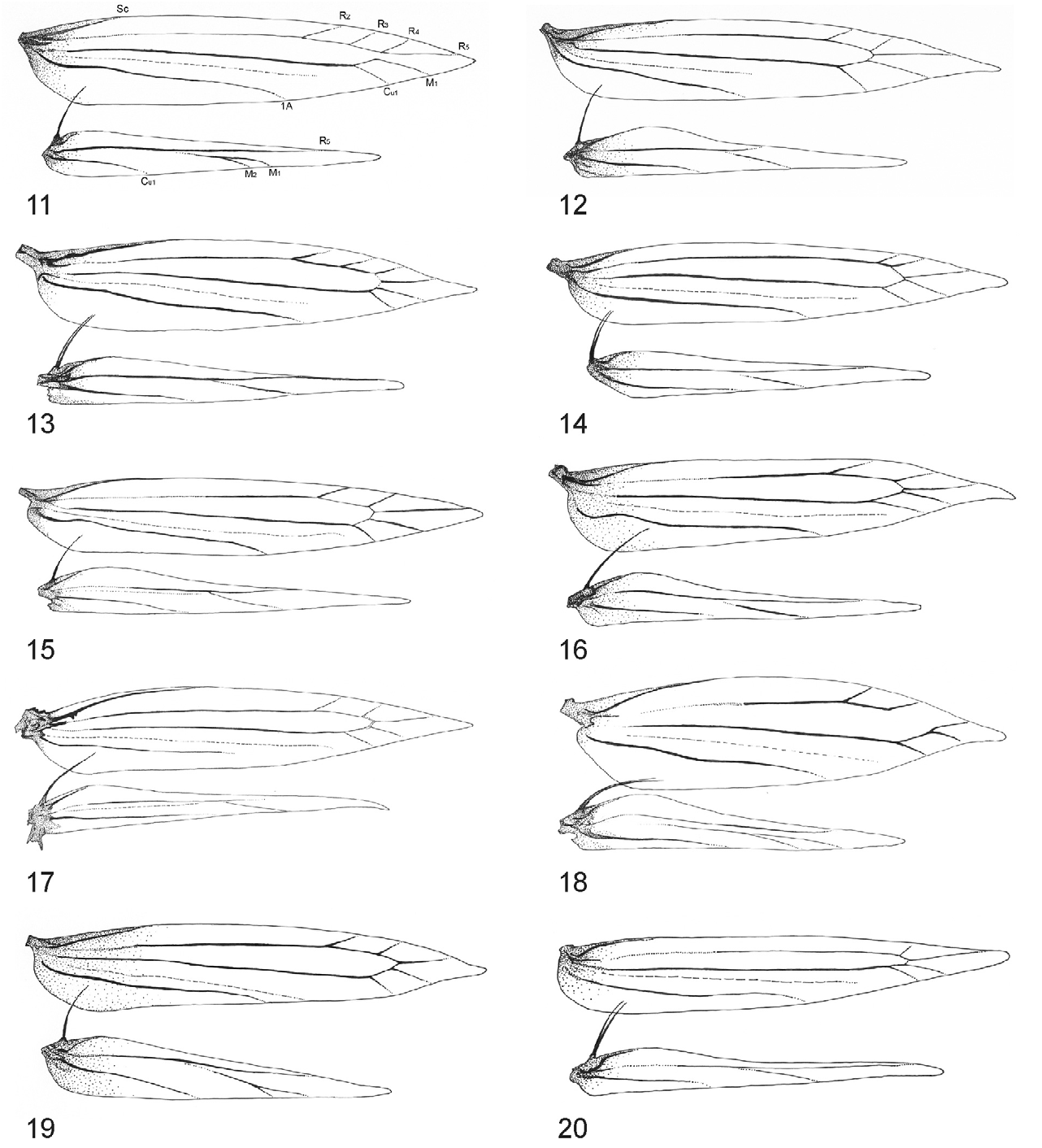
Thorax: Forewing background colour golden-ochreous; apex of forewing broadly rounded; forewing bearing sparsely distributed distinct, mostly fasciate and one strigulate (the basal) white markings; apical marking rounded. Margin of markings distincly black basally with light shading, brownish-greyish apically. Descaled forewing, short lanceolate, elongate, and pointed: maximum width/length ratio ca. 0.2. Forewing venation reduced to 9 veins, apical portion with 6 veins (R 2, R 3, R 4, R 5, M 1, Cu 1); R 2 originates at 4/5 of discal cell; M 1 not stalked, the cell between R 5 and M 1 open, R 5 originates from apex of cell to costa, M 1 arises from apical margin of the discal cell to termen, Cu 1 separate, R indistinct at basal half, CuP indistinct over entire length, 1A thick, separate. Hindwing lanceolate, about 3/4 as long as forewing, with 6 veins: Sc very short, Rs long, running to apical 1/3 of costa, M branched to M 1 and M 2, basal 2/3 of M 1 indistinct, parallel to Rs, Cu 1 absent, Cu 2 thick, simple; A 1 vestigial ( Fig. 11 View FIGURES 11–20 ). Retinaculum as a small fold on Sc. Frenulum in male as a single stout bristle; frenula in female as 2 tightly appressed bristles. Legs slender, with darker rings; fore- and mid- tibia smooth, hind tibia with loosely appressed hairs on its lower surface, hind tarsus smooth, slender and ca. 2× as long as tibia.
Abdomen. Margins of abdominal opening narrowly sclerotized, slightly broadening towards S2, with sclerotized margination of abdominal opening weakly connected on T2 and unconnected on S2; S2 apodemes rather short, ending before opening, slender, with enlarged subbases, slender distally.
Male genitalia. Tegumen long, subconical, with narrow sclerotized arms, flexible; subapex of tegumen covered with short slender appressed setae, apex of tegumen with 5–6 long, stiff setae. Valvae symmetrical, very large and broad, long, bifurcated at apical part; ventral part of valva with 2 thick, sclerotized sutures from base of valva, anastomosing in apical 1/3, ventral surface of valva without setae. Vinculum rather small, caudal portion protruding into a narrow, slender saccus. Transtilla incomplete. Aedoeagus longer than valva, consisting of two distinct morphological parts: (1) narrow tubular, (2) very large bulbous coecum. Anellus strongly developed, sometimes covered with long tufted setae, juxta developed.
Female genitalia. Unknown.
Biology. Unknown.
Distribution. Moths of the Afrotropical Hyloconis occur in the primary rain forests with Fabaceae understore vegetation in Central Africa.
Relationships to other genera. The Hyloconis species sampled here suggest that the genus may be paraphyletic with respect to Neolithocolletis ( Fig. 4 View FIGURE 4 ). While Hyloconis and Neolithocolletis may be synonyms, at this stage we do not formally propose any taxonomic changes and retain both genera until the preimaginal stages of Hyloconis are discovered and additional species are sampled.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Hyloconis Kumata, 1963
| Prins, Jurate De & Kawahara, Akito Y. 2012 |
Hyloconis
| Kumata, T. 1963: 28 |