
Cladotanytarsus (Cladotanytarsus) saetheri, Puchalski & Paasivirta & Giłka, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4394.3.8 |
|
publication LSID |
lsid:zoobank.org:pub:EF77EA75-2056-41E1-B1DF-4BDAD1749621 |
|
DOI |
https://doi.org/10.5281/zenodo.6485429 |
|
persistent identifier |
https://treatment.plazi.org/id/03AC87F2-010C-FFB4-FF6F-FCDFE7ADFE0B |
|
treatment provided by |
Plazi |
|
scientific name |
Cladotanytarsus (Cladotanytarsus) saetheri |
| status |
sp. nov. |
Cladotanytarsus (Cladotanytarsus) saetheri View in CoL , sp. nov.
Figs 1A, B View FIGURE1 ; 2; 3A–E; 4A; 5A–D
Type material. Holotype, adult male: NORWAY , FINNMARK: Helsegård near Karasjok ( 69°26'13"N / 25°41'01"E), 3 August 2003, swarms near river, sweep net, W. Giłka. Paratypes. 47 males as holotype; FINLAND GoogleMaps , OSTROBOTHNIA KAJANENSIS: Oulujärvi-Paltaniemi , 12 June 2017, netting water surface, 3 males, L. Paasivirta ; RUSSIA, FAR EAST: Amur obl., Sigikta River below bridge of the road Neryungri–Never , 3 August 2006, 1 male, T.M. Tiunova ; Magadan obl., Taui River , 26 July 2002, 1 male, E. Khamenkova ; Primorsky Krai, Ussuri River near bridge of the highway Vladivostok – Khabarovsk, 4 August 2003, 1 male, O.V . Orel. CANADA , MANITOBA: Lake Winnipeg: Beaver Creek, 8 June 1971, emergence trap, 2 males, 24 June 1971, emergence trap, 1 male, 22 July 1971, emergence trap, 1 male, 28 July 1971, emergence trap, 1 male, 29 July 1971, emergence trap, 1 male, E. Johnson, M.P. McLean et al.; Gull Harbour, 16 July 1969, light trap, 1 male, P.S.S. Chang; McCreary Island , 25 July 1969, light trap, 1 male, P.S.S. Chang; Victoria Beach, 9 July 1969, light trap, 2 males, P.S.S. Chang (prep. O.A. Saether); USA , COLORADO: Larimer Co., Cache La Poudre River, 15 km NW of Laporte, 26 August 1968, 1 male, F.G. Andrews; Pueblo Co. , Arkansas River / Pueblo Reservoir at: Hobson Ranch, 19 September 1985, 2 males, Pueblo Boulevard Bridge , 15 August 1985, 1 male, Stilling Basin Bridge , 15 August 1985, 2 males, S.J. Herrmann ; MICHIGAN: Newaygo Co., White River near Ramona , 21 May 1982, 2 males, P.C. Baumann & M. Clavla ; SOUTH CAROLINA: Pickens Co., Clemson, Wildcat Creek , 31 March 1976, 1 male, P. Hudson ; WISCONSIN: Burnett Co., 20 km E & 7 km S of Siren , 6 August 1966, 2 male hypopygia, D.C. Hansen.
Derivatio nominis. The specific name is a patronym commemorating the Chironomidae taxonomist and ecologist, Ole Anton Saether ( 1936–2013).
Diagnosis. Acrostichal setae sparse, placed on top of scutum. Wing vein M3+4 ending only slightly proximally of R4+5 or under R4+5. Phallapodeme strongly curved in anterior section. Anal point stocky, with large spinulae densely arranged between prominent crests. Superior volsella with well-developed apical lip and small field of microtrichia confined to dorsolateral surface at base; digitus evenly tapering toward blunt tip. Stem of median volsella strongly elongated, straight or slightly and evenly curved at most, bearing furcate lamellae on apex. Inferior volsella with dorsal lobe forming a nose.
Description. Adult male (n = 73 specimens + 2 male hypopygia).
Colouration (in alcohol). Eyes black. Antenna, tentorium, scutal stripes, scutellum, postnotum and sternum brown to dark brown. Head capsule, mouthparts, ground colour of thorax, legs and abdomen including hypopygium brown or light brown with greenish undertone. Wing and haltere pale brownish.
Head. Eyes reniform, broadly separated by frons. Antenna with 13 distinct flagellomeres; plume fullydeveloped or weak, AR 0.69–0.92 (0.80, n = 18) in specimens with plume fully-developed or AR 0.54–0.67 (0.60, n = 3) in specimens with plume reduced. Frontal tubercles minute, usually in a form of tiny swellings, rarely conical, 8 µm long at most. Lengths of palpomeres 2–5 (in µm) in specimens with antennal plume fully-developed (n = 21): 32–44 (36), 72–100 (88), 79–120 (94), 123–172 (144); palps shorter in specimens with plume reduced (n = 3): 32–40 (36), 66–92 (77), 76–100 (88), 112; pm4> pm3 (n = 21) or pm4 ≤ pm3 (n = 3). Clypeus with 7–14 setae.
Thorax chaetotaxy. Ac 2–6 placed on top of scutum; Dc 5–9 on each side; Pa 1–2 exceptionally 3 on each side (n = 1); Scts 2–4, rarely 6 (n = 2).
Wing ( Fig. 1A, B View FIGURE1 ). Length 1195–1590 (1405) µm. Venation pattern and chaetotaxy slightly variable (also in specimens of the same sample). Veins ending as follows (in order from base to tip): R1, Cu1 (or Cu1 ending under R1), M2+3, M3+4, R4+5 (or M3+4 ending under R4+5), M1+2; VR Cu 1.22–1.43 (1.31). Macrotrichia on C, R, distal half of M1+2, rarely on R1, distal part of R4+5 and distal half of false vein above M3+4, other veins bare; membrane with macrotrichia on r4+5, sometimes in m1+2, exceptionally a couple of macrotrichia in m3+4 (n = 1), other cells bare.
Legs. Fore leg tibia with slightly curved spur 15–24 (20) µm long. Combs of mid and hind leg tibiae separated, ca. twice shorter than spurs; spurs of mid and hind leg unequal in shape (one shorter and straight, second longer and curved) and length: 12–24 µm long on mid leg and 20–36 µm long on hind leg. Basitarsus of mid leg with 2–3 sensilla chaetica. Lengths of leg segments and leg ratios distinctly different in specimens with fully-developed and reduced antennal plume, as shown in Table 1.
fe ti ta1 ta2 ta3 ta4 ta5 LR
p1 465 – 660 265 – 385 550 – 735 295 – 385 220 – 310 150 – 220 85 – 110 1.81 – 2.30 (575) (325) (640) (345) (270) (185) (100) (1.98) 570 – 745 385 – 450 570 – 600 285 195 – 235 145 – 150 105 – 115 1.43 – 1.49 (645) (420)
p2 545 – 690 435 – 565 220 – 330 135 – 195 100 – 180 65 – 115 55 – 85 0.50 – 0.62 (600) (500) (265) (155) (115) (80) (65) (0.53) 610 – 640 495 – 510 250 135 115 85 85 0.49
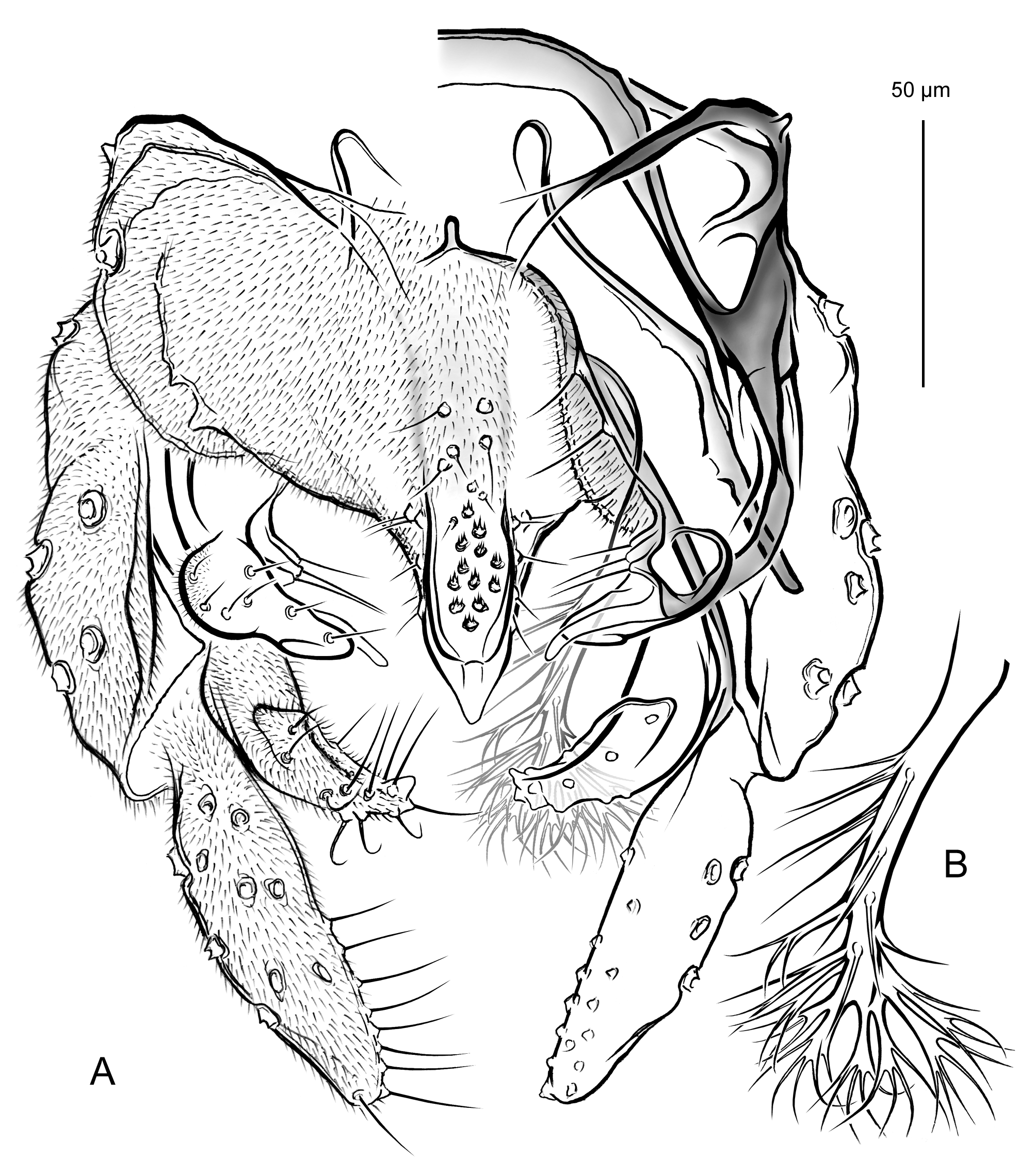
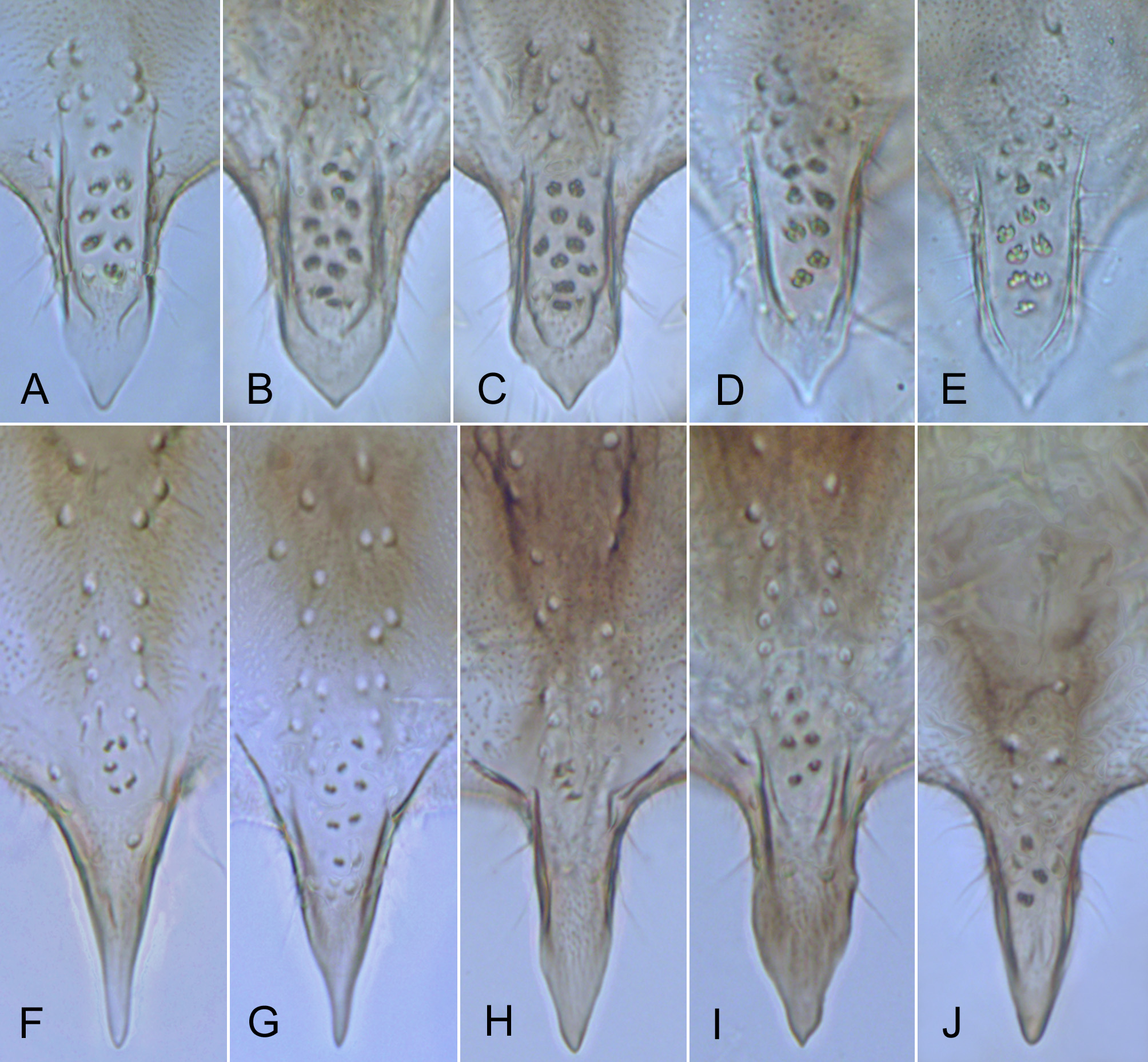
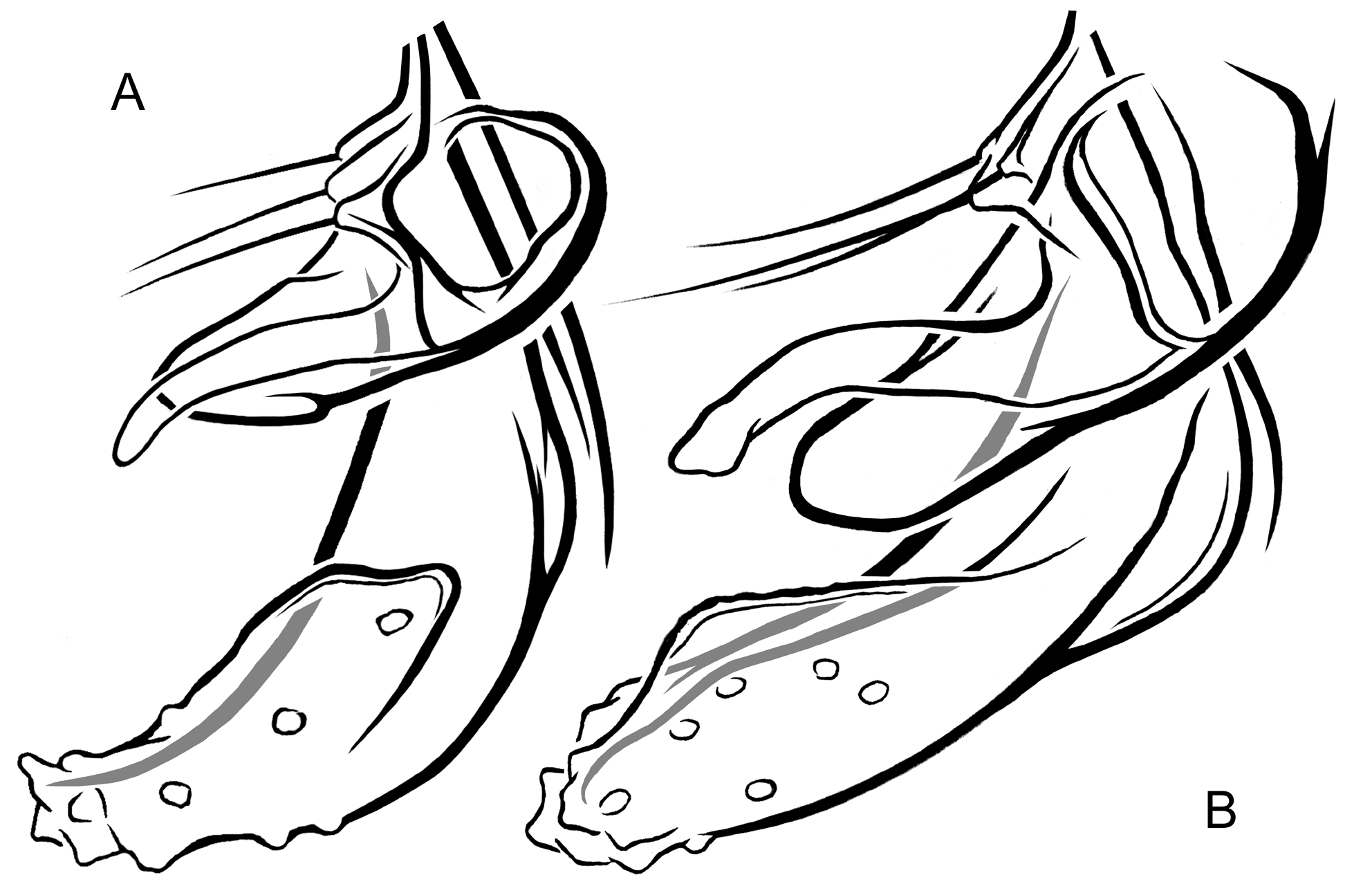
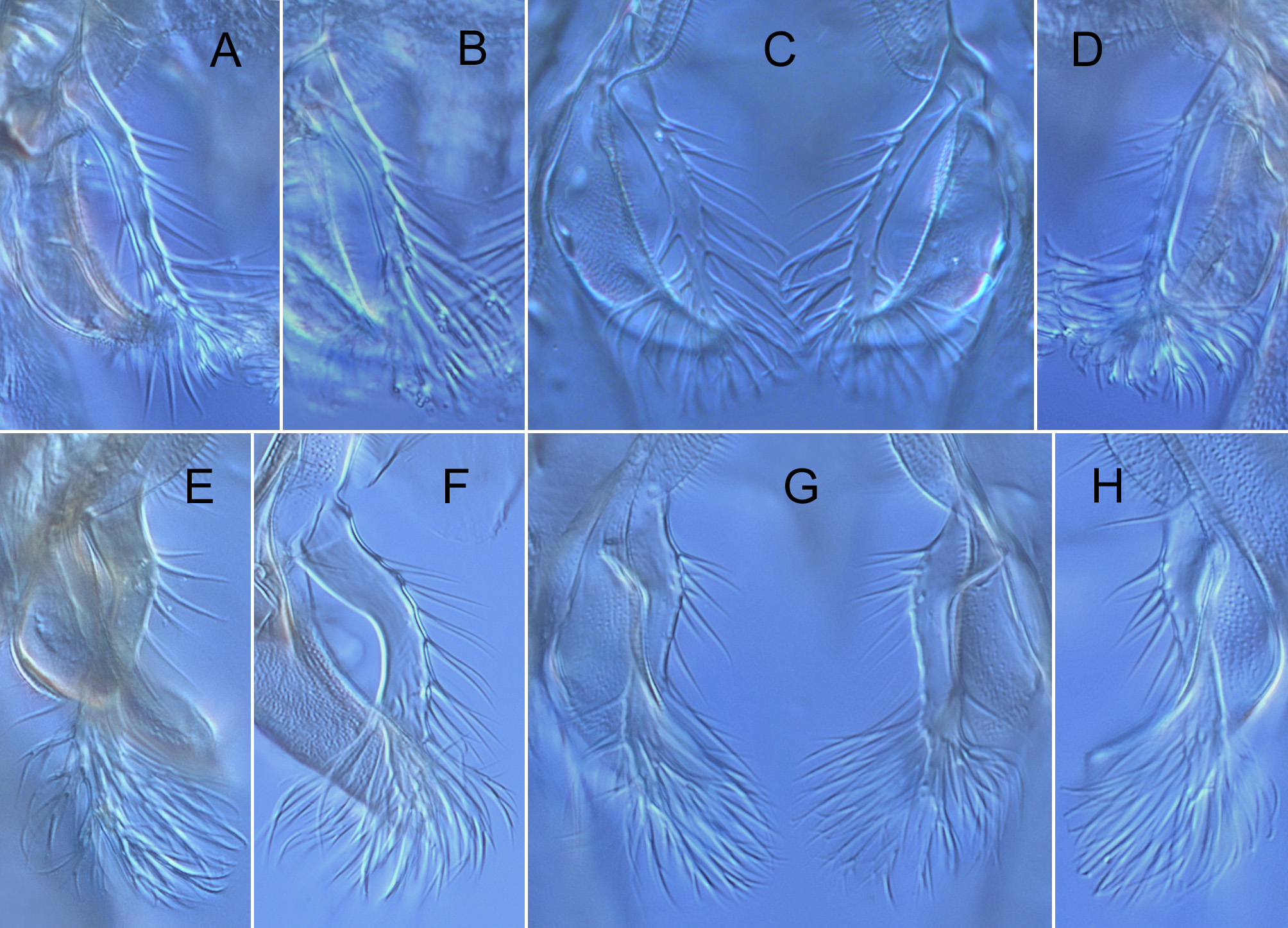
p3 550 – 780 525 – 765 355 – 460 235 – 295 205 – 240 130 – 165 85 – 105 0.56 – 0.65 (675) (660) (405) (260) (225) (150) (95) (0.61) 675-690 645 – 675 330 205 190 120 90 0.49 Hypopygium ( Figs 2 View FIGURE 2 ; 3A–E; 4A; 5A–D). Gonostylus shorter than gonocoxite, 70–100 (80) µm long, rarely longer, up to 120 µm (n = 3, Finnish specimens). Phallapodeme strongly curved in anterior section. Anal tergite with bands of V-type separated, bearing 1 lateral seta on each side and 3–15 (8) median setae arranged irregularly at base of anal point ( Figs 2A View FIGURE 2 ). Anal point stocky, broad at base, tongue-shaped or lanceolate, tapering toward blunt tip or with short nipple-like apical elongation, bearing 6–13 relatively large spinulae (exceptionally 3 spinulae present, n = 1) densely arranged between prominent crests ( Figs 2A View FIGURE 2 , 3A–E View FIGURE 3 ). Superior volsella rounded at base, narrowed at mid length, usually slightly swollen distally, with well-developed apical lip, 4–11 (usually 6–7) dorsal setae and small field of microtrichia confined to dorsolateral surface at base; digitus protruding slightly beyond apex of superior volsella, evenly tapering toward blunt tip ( Figs 2A View FIGURE 2 , 4A View FIGURE 4 ). Stem of median volsella strongly elongated, 40–60 (55) µm long, straight or slightly and evenly curved at most, bearing several setiform and 5–7 rarely 8 furcate lamellae on apex ( Figs 2A, B View FIGURE 2 , 5A–D View FIGURE 5 ). Inferior volsella with slight knee-like extension at base and distinct dorsal lobe forming a nose at mid length of inferior volsella ( Figs 2A View FIGURE 2 , 4A View FIGURE 4 ).
Remarks. Our studies on Cladotanytarsus , by now carried out independently as based on materials from North America (M.P., W.G.) and Fennoscandia (L.P., W.G.) indicated that the same unknown species may occur in both the regions studied. Further materials from the Russian Far East confirmed its wide Holarctic distribution. As a result, Cladotanytarsus saetheri is described and compared with its relative, C. gedanicus . These two species were previously misidentified (Giłka 2009) due to morphological similarities in the hypopygial structure, i.a. peculiar shape of the median volsellae consisted of a long stem bearing dense furcate lamellae on apex ( cf. Giłka 2001: fig. 2). A detailed character analysis indicated several distinct differences between C. saetheri and C. gedanicus , as shown in Figures 1 View FIGURE1 & 3–5 View FIGURE 3 View FIGURE 4 View FIGURE 5 and Table 2.
Though the weakly plumose and abbreviated antennae, shortened palps and changed proportions of legs segments in several examined males of Cladotanytarsus saetheri may disrupt the species definition, these characters were found as a result of a limited flying ability in specimens collected from the water surface (material from Finland). Thus, they are treated separately in the above description; nevertheless, all the presently examined individuals are defined as conspecific. Interestingly, the Finnish specimens were not sampled by simultaneous netting from the air and vegetation. So, it is probable, that they were copulating on the water surface. Such nontypical mode of life may have a diverse background, being e.g. an adaptation to austere conditions, which in a longterm period may lead to behavioural and spatial isolation ( e.g. Serra-Tosio 1974, Hermann et al. 1987, Giłka & Paasivirta 2009, Giłka et al. 2013). This phenomenon usually results in parallel morphological modifications of the wings, antennae, palps and legs, in some cases called as atrophied aberrant characters, and may occur in different Chironomidae groups, including the Tanytarsini ( e.g. Saether 1971, Cranston 1980, Giłka 2011b). Some peculiar leg structures/characters in Tanytarsini , however, are recogniZed as distinct apomorphies ( Giłka 2011b; ZakrZewska et al., in press).
Cladotanytarsus saetheri seems to prefer large, slow flowing rivers and the open shores of large oligotrophic lakes.

| FAR |
Kharazmi University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Chironominae |
|
Tribe |
Tanytarsini |
|
Genus |