
Goliathus orientalis Moser, 1909
|
publication ID |
https://doi.org/10.1080/00222933.2012.763052 |
|
persistent identifier |
https://treatment.plazi.org/id/03AA4B15-1D5D-FFB0-FE7D-C900FE18FA7B |
|
treatment provided by |
Felipe |
|
scientific name |
Goliathus orientalis Moser, 1909 |
| status |
|
Goliathus orientalis Moser, 1909
Goliathus giganteus orientalis Moser, 1909 ; original description by Moser (1909, p. 238); type locality Lindi, Deutsch-Ostafrika; holotype male illustrated by Moser (1909, p. 238) and Sjöstedt (1927b: t.12, f.4).
Goliathus meleagris Sjöstedt, 1927a ; original description by Sjöstedt (1927a, p. 123); type locality Katanga, Congo Belge; holotype male illustrated by Sjöstedt (1927b: t.7, f.1).
Goliathus meleagris ab. conjugatus Sjöstedt, 1927b; original description by Sjöstedt (1927b, p. 9); type locality Katinda, Katanga, Congo Belge; holotype male illustrated by Sjöstedt (1927b, t.10, f.2).
Goliathus meleagris ab. divergens Sjöstedt, 1927b; original description by Sjöstedt (1927b, p. 11); type locality Katinda, Katanga, Congo Belge; holotype male illustrated by Sjöstedt (1927b, t.12, f.3).
Goliathus meleagris ab. maculatus Sjöstedt, 1927b; original description by Sjöstedt (1927b, p. 10); type locality Kinda, Katanga, Congo Belge; holotype male illustrated by Sjöstedt (1927b, t.10, f.4).
Goliathus meleagris ab. niveus Sjöstedt, 1927b; original description by Sjöstedt (1927b, p. 10); type locality Katinda, Katanga, Congo Belge; holotype male illustrated by Sjöstedt (1927b, t.11, f.4).
Goliathus meleagris ab. nyassae Sjöstedt, 1927b; original description by Sjöstedt (1927b, p. 9); type locality Lake Nyassa; holotype male illustrated by Sjöstedt (1927b, t.10, f.3).
Goliathus meleagris ab. oculatus Sjöstedt, 1927b; original description by Sjöstedt (1927b, p. 10); type locality Katinda, Katanga, Congo Belge; holotype male illustrated by Sjöstedt (1927b, t.11, f.1).
Goliathus meleagris ab. orientalis Moser, 1909 ; new combination by Sjöstedt (1927b, p. 17).
Goliathus meleagris ab. partitus Sjöstedt, 1927b; original description by Sjöstedt (1927b, p. 9); type locality Katinda, Katanga, Congo Belge; holotype male illustrated by Sjöstedt (1927b: t.9, f.3).
Goliathus meleagris ab. pustulatus Sjöstedt, 1927b; original description by Sjöstedt (1927b, p. 10–11); type locality Katinda, Katanga, Congo Belge; holotype male illustrated by Sjöstedt (1927b, t.12, f.1).
Goliathus meleagris ab. pustuliferus Sjöstedt, 1927b; original description by Sjöstedt (1927b, p. 11); type locality Katinda, Katanga, Congo Belge; holotype male illustrated by Sjöstedt (1927b, t.12, f.2).
Goliathus meleagris ab. quadrangulatus Sjöstedt, 1927b; original description by Sjöstedt (1927b, p. 10); type locality Katinda, Katanga, Congo Belge; holotype male illustrated by Sjöstedt (1927b, t.11, f.2).
Goliathus meleagris ab. varians Sjöstedt, 1927b; original description by Sjöstedt (1927b, p. 9); type locality unknown, holotype specimen without locality label; holotype male illustrated by Sjöstedt (1927b, t.10, f.1).
Goliathus goliathus usambarensis Preiss, 1933 ; original description by Preiss (1933, p. 89–90); type locality Usambara, Deutsch Ostafrika; holotype male illustrated by Preiss (1933, t.1, f.3); two paratype males illustrated by Preiss (1933, t.2, f.1, 2).
Goliathus goliathus ab. kindanus Endrödi, 1951; original description by Endrödi (1951, p. 45); type locality Kinda, Katanga, Congo Belge; holotype male illustrated by Endrödi (1951, p. 53).
Goliathus goliathus ab. preissi Endrödi, 1951; original description by Endrödi (1951, p. 45); type locality Usambara, Deutsch Ostafrika; holotype male illustrated by Preiss (1933, t.2, f.1; as a paratype of G. goliathus usambarensis Preiss 1933 ) and by Endrödi (1951, p. 54).
Goliathus goliathus orientalis Moser, 1909 (new combination by Burgeon 1928, p. 77; Basilewsky 1956, p. 30 –31).
Goliathus goliatus orientalis Moser, 1909 (new combination by Wiebes 1968, p. 26).
Goliathus orientalis Moser, 1909 (new status by Lachaume 1983, p. 28 –32).
Goliathus orientalis var. preissi Endrödi, 1951 (new status by Lachaume 1983, p. 28).
Goliathus orientalis var. pustulatus Sjöstedt, 1927b (new status by Lachaume 1983, p. 28).
Goliathus orientalis var. undulatus Lachaume, 1983 , not Kraatz (1897, p. 251); name published by Lachaume (1983, p. 28) but preoccupied by Goliathus giganteus var. undulatus Kraatz 1897 (name published by Kraatz 1897, p. 251).
Goliathus orientalis preissi Endrödi, 1951 (new status by Lachaume 1983, pl.19).
Goliathus orientalis undulatus Lachaume, 1983 , not Kraatz 1897; name published by Lachaume (1983, p. 28) but preoccupied by Goliathus giganteus var. undulatus Kraatz 1897 (name published by Kraatz 1897, p. 251).
Goliathus orientalis pustulatus Sjöstedt, 1927b (new status by Lachaume 1983, pl.21).
Diagnosis, separation from allied species and distribution
Most previous descriptions and diagnoses of G. orientalis have focused on details of the dorsal colour patterns. I have accordingly prepared the following anatomical re-descriptions of both sexes based on the study of 43 male and 24 female specimens from Shaba or Katanga Province in the République Démocratique du Congo (representative specimens illustrated in Figure 1 View Figure 1 ) and seven male and three female specimens from the Usambara Mountains in Tanzania (representative specimens illustrated in Figure 2 View Figure 2 ) .
Male. Length 50–91 mm, exclusive of cephalic horns, which add 3–5 mm to overall body length. Integument black, brownish-black, or greenish-black, strongly shining, except for areas where the surface is covered with white and black patterned markings, which have a matte appearance. Areas with markings include dorsal surface of head, dorsal and ventral surfaces of pronotum, mesoscutellum, elytra and mesepimera. Head elongate, with stout Y-shaped anteriorly directed horn at apex and with a short rectangular projection above each eye. Eyes round, brown or black. Antennae short, small, of typical scarab form with large scape and lamellate terminal three-segmented club. Pronotum broad, approximately oval in shape, except for a small transverse basal projection extending over the mesoscutellum and a small rounded projection extending over the head; frontal angles acute and small, lateral margins broadly rounded, basal angles obtuse and broadly rounded. Disc of pronotum convex, except for small concave areas behind the apical angles and a larger shallow concave area on the anterior portion of the disc. Ventral surface of pronotum smooth, white. Mesoscutellum small, triangular, longer than wide, with a median triangular elevated area, white with a black “V” on disc. Elytra large, trapezoidal in shape, elytral length greater than width across both elytra; each elytron strongly convex with humeral and apical calli distinctly tumescent. Mesosternum finely and distinctly punctate, with sparse elongate orange pubescence. Metasternum smooth medially and finely punctate laterally, with elongate orange pubescence. Meso-metasternal process well developed, forming a stout triangular projection that is densely covered in short orange pubescence along its anterior surface. Abdominal ventrites matte, without distinct punctures, with small areas of short orange pubescence laterally. Pygidium black, transverse, with a fringe of long orange setae along apical margin. Legs stout, massive, strongly shining. Protibiae rectangular, lacking lateral denticles but with well-defined lateral and ventral carinae and a small patch of short orange pubescence along interior margin at base. Apex of protibiae with two stout triangular denticles: an anteriorly directed projection along the lateral margin, and a ventrally directed projection along the interior margin. Protarsi elongate and somewhat enlarged relative to mesotarsi and metatarsi, with individual tarsomeres distinctly longer than wide. Protarsal claws massive, much larger than mesotarsal or metatarsal claws. Mesothoracic and metathoracic legs similar in size and structure: mesofemora and metafemora robust; mesotibiae and metatibiae with a fringe of orange setae along inner margin and with two spurs at apex; mesotarsi and metatarsi stout, tarsomeres 1–4 not much longer than wide, tarsomere 5 elongate, over twice as long as wide with a pair of large claws. Head, dorsal and ventral surface of pronotum, mesoscutellum, mesepimeron and elytra covered in white cretaceous markings; pronotum with six large irregular black longitudinal markings extending from base to apex, black markings covering less than half the pronotal surface; elytron with a network of black markings on disc such that the white markings are reduced to irregular rows or lines of round or ovate marks. In populations from the Usambara Mountains, Tanzania, the black and white elytral markings are often more linear ( Figure 2 View Figure 2 ) than rounded ( Figure 1 View Figure 1 ).
Female. As described for male except: body size somewhat smaller, length 64 to 82 mm. Dorsal surface of head lacking horns, apex of head shovel-shaped, with a small tubercle above each eye. Protibiae with two lateral and two apical denticles; protarsi only slightly more elongate than mesotarsi and metatarsi. Mesotibiae and metatibiae each with a single lateral denticle and multiple apical denticles and spurs; mesotibiae with two short apical denticles and two spurs; metatibiae with four short apical denticles and two spurs. Pronotum more elongate than in male, with a distinct tubercle along anterior margin adjacent to head and a well-defined concave impression on the pronotal disc behind this tubercle. Basal half of pronotum strongly shining, with large well-defined punctures, lacking white markings in many specimens. Mesoscutellum black with small lateral white markings and occasionally a small white median marking as well. Pygidium more elongate than in male, and distinctly triangular.
Separation of adults of G. orientalis from the adults of allied species has not always been straightforward because of the strong similarities in colour pattern between certain rare colour forms of G. goliatus and the common colour forms of G. orientalis . As with other groups of Cetoniinae ( Holm and Marais 1992), the male reproductive structures offer a set of characters that can be helpful in diagnosing species within the genus Goliathus . Sjöstedt (1927b) was the first to study male reproductive structures in Goliathus and proposed full species status for his G. meleagris based on the differences in the structure of the aedeagus. Wiebes (1968) also studied male reproductive structures and based his revision of Goliathus in part on characters of the aedeagus. Given the importance of these structures for species diagnoses in this genus, I have provided photographs of the male aedeagi of G. albosignatus , G. goliatus and G. orientalis in Figure 3 View Figure 3 . In general, the aedeagus of G. albosignatus is smaller and stouter than in the other species of the genus ( Wiebes 1968), whereas the aedeagus of G. orientalis is more elongate and slender than that of G. goliatus ( Figure 3 View Figure 3 ). As illustrated in Figure 3 View Figure 3 , the aedeagus of G. goliatus has a pronounced median constriction and has the apices distinctly rounded, characters that are less strongly pronounced in the more slender and elongate aedeagus of G. orientalis .
For much of its range, G. orientalis is the largest cetoniine scarab so it should be easily recognizable by its massive size and distinctive dorsal colour pattern ( Figures 1 View Figure 1 , 2 View Figure 2 ). In the southern Congo River Basin, G. orientalis overlaps with G. goliatus , which is similar in size but can often be distinguished from G. orientalis on the basis of dorsal colour pattern characteristics. Males of G. goliatus tend to have more extensive black markings on the pronotum, whereas males of G. orientalis tend to have the pronotum predominantly white with black markings. Southern Congo populations of G. goliatus have the elytra predominantly reddish-brown to dark brown in both sexes, occasionally with scattered white markings but generally lacking the large irregular ovate patches seen in G. orientalis . According to Basilewsky (1956), these species also differ in their ecological habitat associations: G. goliatus is usually associated with tropical forests and forest patches, whereas G. orientalis is usually associated with savannah ecosystems. In Tanzania, G. orientalis overlaps with G. albosignatus Boheman, 1857 , another savannah or woodland species, which is generally significantly smaller in size (length in both sexes 35–65 mm), has elytra with transverse or oblique black and white markings, and often has white markings on the pygidium, at least in Tanzanian populations ( Holm and Marais 1992).
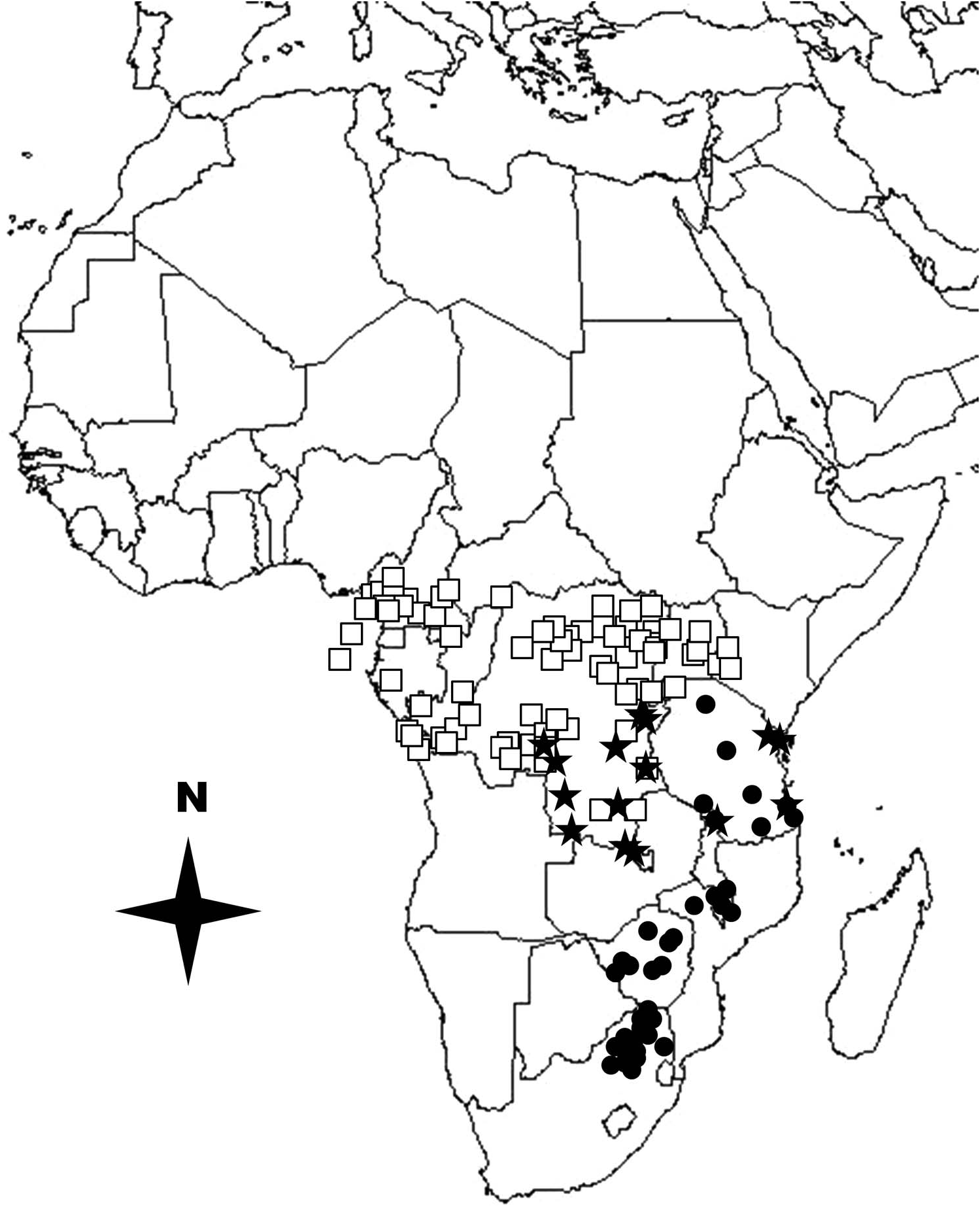
The exact limits of the distribution of G. orientalis are poorly known, but specimens have been collected from Katanga (also known as Shaba) Province in the southern portions of the République Démocratique du Congo, east across central Africa to Lindi and the Usambara Mountains of Tanzania ( Wiebes 1968; Lachaume 1983). In Figure 4 View Figure 4 , I have mapped the known collecting localities for G. albosignatus , G. goliatus and G. orientalis based on museum specimen records and publications by Kraatz (1897, 1898), Moser (1909), Sjöstedt (1927b), Burgeon (1928), Preiss (1933), Endrödi (1951), Basilewsky (1956), Wiebes (1968), Lachaume (1983) and Holm and Marais (1992).
Notes on synonymies
The earliest published name for the species discussed in this paper is Goliathus giganteus orientalis Moser, 1909 . This name has 18 years of priority over the next published name, Goliathus meleagris Sjöstedt, 1927a . Moser’s holotype male was collected at Lindi on the Indian Ocean (at the time in Deutsch Ostafrika, now in modern-day Tanzania), and Sjöstedt’s holotype male was collected in Katanga in the Congo River Basin (at the time in Congo Belge, now in modern-day République Démocratique du Congo). The two type specimens clearly represent a single species ( Wiebes 1968; Lachaume 1983), which by priority ( ICZN 1999; Article 23) is named G. orientalis Moser.
Study of additional material of this species led Lachaume (1983) to conclude that most of the varieties described by Sjöstedt (1927b) and Endrödi (1951) simply represented normal intrapopulational colour variants. Consequently, all but three of these names were treated as straightforward synonyms of G. orientalis by Lachaume (1983) and subsequent authors (e.g. Marais and Holm 1992; Krajčík 1998).
As noted above, Lachaume (1983) created several taxonomic problems in his treatment of G. orientalis . Lachaume placed most of the names proposed by Sjöstedt and Endrödi in synonymy, but also recognized three well-marked colour forms or varieties, which he called G. orientalis var. pustulatus Sjöstedt , G. orientalis var. preissi Endrödi and G. orientalis var. undulatus Kraatz. These names were treated as infrasubspecific taxa in the text of Lachaume’s work, but were labelled as subspecific taxa ( G. o. pustulatus, G. o. preissi and G. o. undulatus) in the plates of this same volume.
Two of these taxa, which Lachaume called G. orientalis var. pustulatus and G. orientalis var. undulatus , represent intrapopulational colour variants that occur in most southern Congo populations of G. orientalis ( Sjöstedt 1927b; Burgeon 1928; Lachaume 1983). Hence, these names should be considered infrasubspecific rather than subspecific, and should be treated as straightforward synonyms of G. orientalis . The attribution of “ var. undulatus ” to Kraatz (1897) is incorrect; in a subsequent publication, Kraatz (1898) indicates that this varietal name was actually based on a specimen from Cameroon, which is outside the known distribution of G. orientalis but a well-known collecting area for G. goliatus ( Wiebes 1968) . The name Goliathus giganteus var. undulatus Kraatz, 1897 , is now considered to be a straightforward synonym of G. goliatus ( Krajčík 1998) .
Lachaume (1983) also identified a third colour form within G. orientalis , which he called G. orientalis var. preissi . He applied this name to a well-defined and regionally restricted colour form from the Usambara Mountains of northeastern Tanzania that has alternating white and black longitudinal markings on the elytra of both sexes. However, the name G. goliathus usambarensis Preiss 1933 is clearly the first available name for this taxon, predating the description of G. goliathus ab. preissi Endrödi 1951 by 18 years. In fact, Endrödi (1951) indicates that his G. goliathus ab. preissi was based on an illustration published by Preiss (1933) of a paratype specimen of G. g. usambarensis Preiss! The name G. orientalis preissi should therefore be considered a junior subjective synonym of G. g. usambarensis.
The taxonomic status of the Usambara Mountain populations of G. orientalis requires additional investigation. During my studies of museum collections, I examined seven males and two females of G. orientalis that had been collected in the Usambara Mountains (representative specimens illustrated in Figure 2 View Figure 2 ). Five of the males and one of the females had elytra that were largely white, with the black markings reduced to irregular longitudinal lines. Similar specimens were illustrated by Preiss (1933; as G. g. usambarensis) and Lachaume (1983; as G. o. preissi). It is possible that these specimens represent a distinct geographic form or subspecies within G. orientalis . However, I also examined three specimens collected in the Usambara Mountains that had elytral patterns similar to specimens collected elsewhere in Tanzania and in the southern Congo River Basin. Further investigations of the taxonomic status of the Usambara Mountain populations of G. orientalis are clearly needed. This is not merely a question of obtaining additional specimens for study from existing institutional or private collections, however. Much of the material of G. orientalis that is currently available for study has been obtained through the commercial specimen trade and lacks some or all pertinent data such as longitude, latitude and elevation of capture, date of collection, associated plant species and associated vegetation communities. Careful de novo collections of georeferenced specimens with associated ecological and environmental data would be most helpful in resolving the status of the Usambara Mountain populations of G. orientalis . If future studies indicate that this cluster of populations is distinct enough to merit recognition at the subspecific level, the correct subspecific name for the taxon would be G. orientalis usambarensis Preiss, 1933 , not G. orientalis preissi Endrödi, 1951 .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Goliathus orientalis Moser, 1909
| Mawdsley, Jonathan R. 2013 |
Goliathus orientalis
| Lachaume G 1983: 28 |
Goliathus orientalis var. preissi Endrödi, 1951
| Lachaume G 1983: 28 |
Goliathus orientalis var. pustulatus Sjöstedt, 1927b
| Lachaume G 1983: 28 |
Goliathus orientalis undulatus
| Lachaume G 1983: 28 |
| Kraatz G 1897: 251 |
Goliathus goliatus orientalis
| Wiebes JT 1968: 26 |
Goliathus goliathus
| Endrodi S 1951: 45 |
| Endrodi S 1951: 53 |
Goliathus goliathus
| Endrodi S 1951: 45 |
| Endrodi S 1951: 54 |
Goliathus goliathus orientalis
| Basilewsky P 1956: 30 |
| Burgeon L 1928: 77 |
Goliathus meleagris Sjöstedt, 1927a
| Sjostedt Y 1927: 123 |
Goliathus meleagris
| Sjostedt Y 1927: 9 |
Goliathus meleagris
| Sjostedt Y 1927: 10 |
Goliathus meleagris
| Sjostedt Y 1927: 17 |
Goliathus orientalis var. undulatus
| Lachaume G 1983: 28 |
| Kraatz G 1897: 251 |
| Kraatz G 1897: 251 |