
Perameles papillon, Travouillon & Phillips, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4378.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:FD162D89-A6D6-44FA-AB5E-9DA724F7B62A |
|
DOI |
https://doi.org/10.5281/zenodo.6485403 |
|
persistent identifier |
https://treatment.plazi.org/id/453893E8-55DC-481A-B095-EAF67B434B8A |
|
taxon LSID |
lsid:zoobank.org:act:453893E8-55DC-481A-B095-EAF67B434B8A |
|
treatment provided by |
Plazi |
|
scientific name |
Perameles papillon |
| status |
sp. nov. |
Perameles papillon sp. nov.
Figs. 5–7 View FIGURE 5 View FIGURE6 View FIGURE 7
Holotype. WAM M571, skin and skull, female.
Type locality. Ooldea , South Australia (SA).
Paratypes. WAM M126, skull, sex unknown, Eucla Pass , Western Australia (WA); WAM M570, skin and skull, female, Ooldea (SA); WAM M572, skin and skull (juvenile), male, Ooldea (SA); WAM M574, skin and skull ( Juvenile ), male, Ooldea (SA); WAM M576, skin and skull, female, Ooldea (SA); WAM M577, skin and skull, female, Ooldea (SA); AM M4352, Skin and skull, female, Rawlinna (WA); AM M31547, alcohol specimen, male, Ooldea (SA) ; BMNH 1925.10 .8.26, skin, male, Ooldea (SA) ; BMNH 1925.10 .8.27, skin, female, Ooldea (SA); SAM M3973, alcohol specimen and skull, female, collected between Ooldea and Tallaringa (SA); SAM M4640, skin and skull, male, Ooldea (SA).
Referred specimens. AMNH 196371, 2 left subfossil dentaries, Horseshoe Cave, Nullarbor (WA); AMNH 220153 View Materials and AMNH 220154 View Materials , juvenile subfossil skulls, south west Loongana , Nullarbor (WA); AM S1821, skull, juvenile, Rawlinna (WA); AM M2986, skin and skull, female, Fisher, Nullarbor (SA); AM M4353, skin and skull, juvenile female, Rawlinna (WA); AM M4850, skin and skull, male, Ooldea (SA); AM M4851 and M4852, skin and skull, female, 9 miles west of Ooldea (SA); AM M3049, alcohol specimen, juvenile female, Ooldea (SA); AM M4368–M4369, alcohol specimen, juvenile female, Rawlinna (WA); AM M4370–M4371, alcohol specimen, juvenile male, Rawlinna (WA); AM M4939 and M4941, alcohol specimen, pouch young male, Ooldea (SA); AM M4940, alcohol specimen, pouch young female, Ooldea (SA); AM M4942–4943, alcohol specimen, male, East West Line (SA); AM M4944–4945, alcohol specimen, female, East West Line (SA); AM M4978–4979, alcohol specimen, pouch young female of M4852, 9 miles west of Ooldea (SA); AM M31545, alcohol specimen, juvenile male, Ooldea (SA); AM M31548, alcohol specimen, male, Ooldea ? (SA); MAGNT U7605, skull, specimen bred in Adelaide in 1926 (original locality unknown); NMV C7173 and C7174, subfossil skulls, N11 Cave, Nullarbor East (SA); NMV C7175 and C7176, subfossil skulls, Weebubbie Cave, Nullarbor (WA); NMV C7177 and C7178, subfossil skulls, Weekes Cave, Nullarbor (SA); NMV C7180, 2 subfossil dentaries, Koonalda Cave, Nullarbor (SA); QM J5028, alcohol specimen, female, Nullarbor (SA); SAM M846, alcohol specimen and skull, juvenile female, Ooldea (SA); SAM M1394, mount, male, Ooldea (SA); SAM M1395, mount, Ooldea (SA); SAM M1396, mount, female, Ooldea (SA); SAM M1397, skin, male Ooldea (SA); SAM M1398, skin, female Ooldea (SA); SAM M3926–3927, M3976, M3981–3982, M3984, M3987–3988, alcohol specimen and skull, female, no location data; SAM M3977, M3983, M3985–3986, alcohol specimen, juvenile, no location data; SAM M3980, M3989, alcohol specimen and skull, male, no location data; SAM M4639, skin and skull, juvenile female, Ooldea (SA); SAM M5226, mount, no data; SAM M6302, many subfossil skulls, Weebubbie Cave, Nullarbor (WA); SAM M6303, many subfossil skulls, Weekes Cave, Nullarbor (SA); SAM M6304, many subfossil skulls, N11 Cave, Nullarbor East (SA); SAM M6305, many subfossil skulls, Koonalda Cave, Nullarbor (SA) ; WAM 76.4.21, skull, Abrakurrie Cave, Nullarbor (WA); WAM 67.4.114–116, 67.10.305–307, skulls, Homestead Cave, Nullarbor (WA); WAM 66.6.45–53, 66.1.54, 68.3.63–67, 69.5.17, skulls, Murra-el-elevyn Cave, Nullarbor (WA); WAM 69.7.557– 559, skulls, Kjeldahl Cave, Cocklebiddy Roadhouse, Nullarbor (WA); WAM 73.1.107, 74.5.39, skulls, Webbs Cave, Nullarbor (WA); WAM 67.5.128, 67.5.130, skulls, Warbla Cave, Nullarbor (SA); WAM 67.10.420–421, 68.3.118, skulls, Blowhole, Eucla, Nullarbor (WA); WAM 67.4.250, 72.1.8, 72.1.1088–1089, skulls, Horseshoe Cave, Nullarbor (WA); WAM 66.6.45–53, skulls, Graham’s Cave, Nullarbor (WA); WAM 68.3 View Materials .51, skull, cave 3 3/ 4 mile South of Cocklebiddy Cave , Nullarbor (WA) ; WAM 68.3.33, skull, Mullamullang Cave, Nullarbor (WA); WAM 67.11.32, skull, Capstan Cave, Nullarbor (WA); WAM 67.4 View Materials .343, skull, cave 23 miles west of Eucla , Nullarbor (WA) ; WAM 68.3 View Materials .20, skull, cave 6 miles south of Madura , Nullarbor (WA) ; WAM 67.9 View Materials .134, cave near Madura , in doline, Dingo Douga, Nullarbor (WA); WAM 66.3.14–16, 67.10.177–182, skulls, Firestick Cave, Nullarbor (WA); Nullarbor (WA).
Diagnosis. Perameles papillon sp. nov. differs from other species of Perameles in having a dark horizontal bar on its ears, a bracelet of light brown fur around its wrists and covering the heel of the foot and the posterior edges of the footpad, a dark butterfly pattern on its rump, shorter pes and tail (see results), enlarged lacrimal crest (except for P. gunnii ), enlarged bullae (largest of any Perameles ), accessory fenestrae extending anteriorly to the canine, maxillopalatine fenestrae extending posteriorly to the M3, well-developed squamosal epitympanic sinus. Perameles papillon sp. nov. differs from all members of Peramelemorphia (except P. myosuros ) in having females distinctively larger than males.
Etymology. Papillon, French for butterfly, in reference to the butterfly shaped pattern on its rump.
Remarks. Westerman et al. (2012) subsampled specimen AM M31545 from the Australian Museum, which was attributed to Perameles b. notina , from Ooldea in South Australia. After scoring specimens from this region, including the rest of the Nullarbor Plain in Western Australia, and comparing them to the holotype of Perameles b. notina (then named Perameles myosura notina ) BMNH 43.8.12.21 from the head of St Vincent Gulf in South Australia, it became evident that they did not match in morphology, the type being much larger and several features on the skin and skull did not match any of the specimens recovered from the Nullarbor region. As a result, AM M31545 and all specimens found in the Nullarbor regions (present in collections in various museums) are considered here a new species.
The results of our analysis also show that Perameles papillon sp. nov. is distinct morphologically from Perameles bougainville and P. eremiana , but also distinct to the currently unrecognised P. myosuros , P. notina and P. fasciata . Our results further suggest that the latter three taxa are distinct taxa and should no longer be subsumed in P. bougainville (see discussion below).
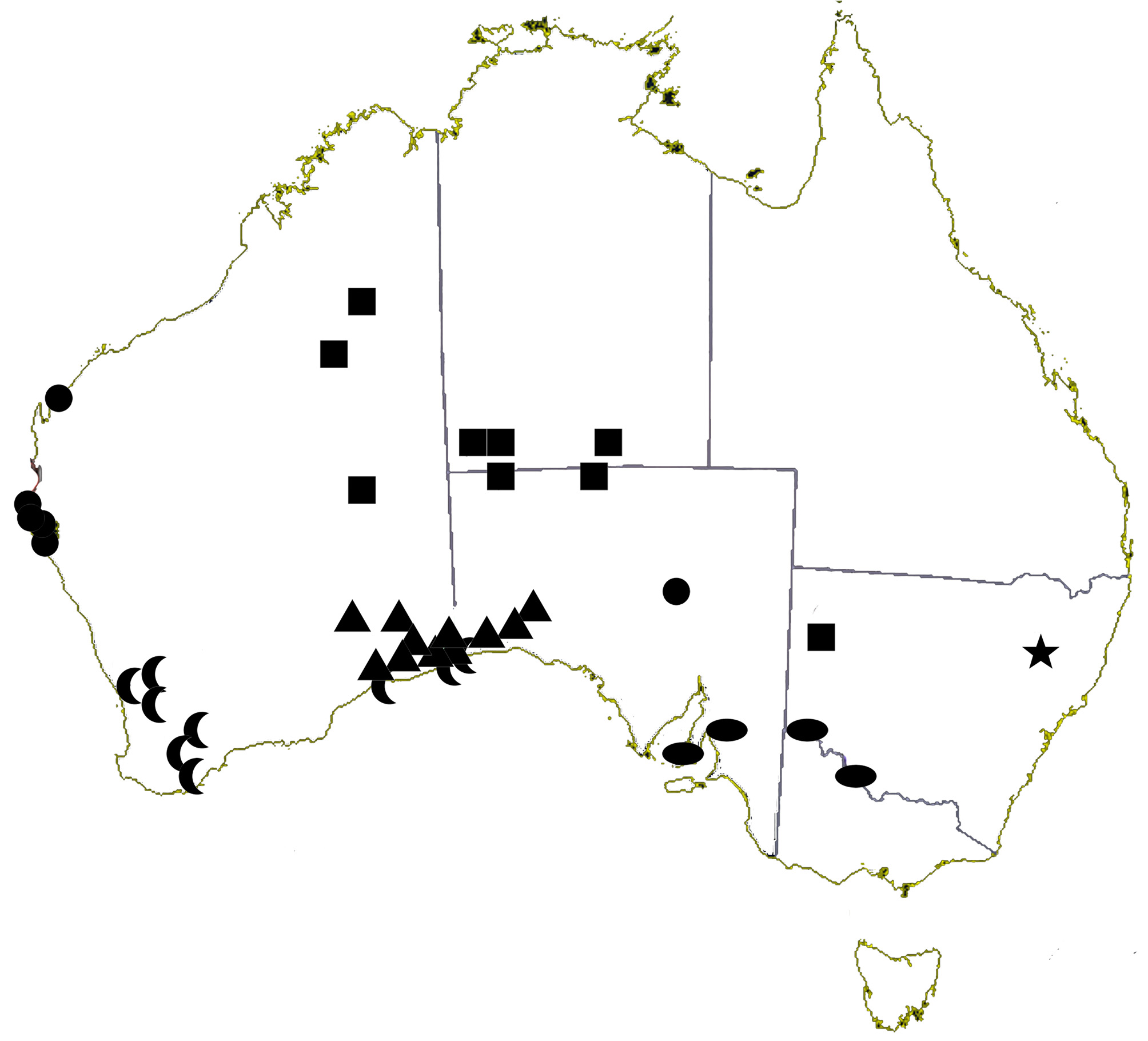
Distribution. Collected only in the Nullarbor Plain, on either side of the Western Australian and South Australian border (triangles in Fig. 8 View FIGURE 8 ).
Description. The description of the skin is a comparison of Perameles papillon sp. nov., represented by Holotype WAM M571 ( Fig. 5 View FIGURE 5 ), to P. b. bougainville , represented by WAM M3641 (Suppl. Fig. 2 View FIGURE 2 ), and P. b. notina , represented by the holotype BMNH 43.8.12.21, unless specified otherwise (Suppl. Fig. 3 View FIGURE3 ). P. b. fasciata and P. b. myosuros are not used in the comparison here as they were not sampled by Westerman et al. (2012), though we do suspect they are distinct taxa and will be investigated in future research.
P. papillon sp. nov. is similar in size to P. b. bougainville but much smaller than P. b. notina , showing no marked sexual dimorphism between males and females, though females seem to be a little larger (see Table S2 –3 for measurements, Fig. 3 View FIGURE3 for morphometric analysis). The fur on the dorsal side of the head is a little darker and browner than P. b. bougainville , but lighter than P. b. notina . The ventral side is cream in colour as in P. b. bougainville whereas it is greyer in P. b. notina . The ears have a noticeable dark horizontal bar across them, absent in both P. b. bougainville and P. b. notina or any other known peramelemorphian. Note that none of the wet specimens of P. papillon sp. nov. show this feature (dark bar on ear) but it is present in all dry skins, and this is suspected to be as a result of accelerated fading in alcohol specimens. The ears are long as is P. b. bougainville , but shorter than P. b. notina . There are long dark vibrissae present at the front of the snout, above the eye and on the cheek below the eye. These are also present in P. b. bougainville and P. b. notina , but less obvious in P. b. bougainville . The body is also darker and browner on the dorsal side than in P. b. bougainville but lighter than P. b. notina . Both P. papillon sp. nov. and P. b. bougainville are cream coloured on the ventral side, while P. b. notina is mostly grey except for patches of cream. A complex pattern of different coloured bars is present on the rump (juveniles WAM M574 and M572 show the early stages of the development of this pattern on the rump), which is absent in P. b. bougainville (P. b. bougainville has a single very faint darkish bar across its rump that tapers laterally) but present in P. b. notina , though the holotype is faded, making this pattern less obvious. The pattern resembles that of P. gunnii and P. b. notina but it is less well defined. There isn’t a central dark dorsal bar as in P. gunnii , and most likely in P. b. notina , and the light bars are bicoloured, cream and golden brown (golden brown only in P. gunnii and P. b. notina ). The anterior most two dark bars are almost completely black or at least dark brown (specimens are likely faded), and looks like butterfly wings in shape in dorsal view. The posterior darkest bars are much lighter in colour and less well defined, distinguishing P. papillon sp. nov. from P. gunnii and P. b. notina , which have a well-defined posterior bar. The tail is short as in P. b. bougainville (longer in P. b. notina ), but it is tricoloured (dark brown dorsally, beige laterally and cream ventrally) instead of bicoloured as in P. b. bougainville (the tail is dark brown dorsally and cream ventrally) or unicoloured as in P. b. notina (the tail is dark grey). The forelimbs are light brown in colour on the dorsal side up to the wrist, where it surrounds the wrist like a bracelet. The manus and the ventral side of the forelimbs are cream in colour. This pattern is different to P. b. bougainville , which lacks the bracelet of brown fur around the wrist, and is lighter in colour dorsally. The forelimbs of P. b. notina are completely grey. The manus has the three large toes (digit II, III and IV), and the typically reduced digits I and V as in all Peramelidae . Compared to P. b. bougainville and P. b. notina , the digits are shorter but have more pronounced toepads as in P. b. notina (the toes of P. b. bougainville more slender). The hindlimbs, in the region of the tibia, are cream in colour anteriorly and greyish brown on the posterior side. The darker colour invades the heel of the cream coloured pes, and the posterior edges of the footpad, unlike in P. b. bougainville where the pes is completely cream in colour and P. b. notina where the pes is completely grey in colour. The footpad is hairless posterior to digit I to the heel as in P. b. notina , while in P. b. bougainville , this area is covered in fur. As in the manus, the pes has shorter toes with thicker toepads than P. b. bougainville and P. b. notina .
The description of the skull and teeth is based on paratype WAM M577 for Perameles papillon sp. nov. ( Fig. 6 View FIGURE6 ), and describes only features that differ from other Perameles , using WAM M6166 for P. b. bougainville , BMNH 43.8.12.21 for P. b. notina , WAM M2629 for P. eremiana , WAM M16590 for P. gunnii (Suppl. Fig. 4 View FIGURE4 ), QM JM18575 for P. pallescens and WAM M44421 View Materials for P. nasuta (see Travouillon 2016a), unless specified otherwise. The skull of P. papillon sp. nov. is similar in size to P. b. bougainville and P. eremiana (much smaller than P. b. notina , P. gunnii , P. pallescens and P. nasuta ) but it is shorter than P. eremiana (nasals shorter in length), and much wider than both P. b. bougainville and P. eremiana . In dorsal view, the lacrimal crest is highly developed, as seen in P. gunnii and P. b. notina , and is visible as a distinct posteriorly orientated process in the orbit. The lacrimal crest is less developed in all other species of Perameles . No sagittal crest is present, as in all Perameles except in males of P. nasuta and P. pallescens . In ventral and lateral view, the alisphenoid tympanic process, or bullae, is highly distinctive in being the most developed (the second largest bullae) within the genus. P. b. bougainville has the smallest bullae, followed by P. nasuta , P. gunnii , P. eremiana , P. papillon sp. nov. and P. b. notina , in increasing order of size. On the palate, accessory fenestrae are present between the incisive fenestrae and the maxillopalatine fenestrae as in all Perameles except P. nasuta and P. pallescens , but they extend anteriorly to the canine, unlike other species of Perameles except P. b. notina where the anterior extension of the accessory fenestrae is posterior to the canines. The maxillopalatine fenestrae also extend further posteriorly than in other Perameles except P. b. notina , from the anterior of the M3 to level with the middle of M3, instead of level with M2. The dividing septa of the maxillopalatine fenestrae are thick anteriorly and posteriorly, thinning centrally, as in Perameles nasuta (very thin in all other Perameles ). Palatine fenestrae are present and large as in P. b. bougainville (broken in P. b. notina ). In lateral view the orbitosphenoid is identified as a very reduced bone, as in P. nasuta , P. pallescens and P. gunnii (much larger in P. b. bougainville and P. eremiana , broken in P. b. notina ). The antorbital fossa is very deep as in all Perameles except in P. nasuta and P. pallescens . The squamosal epitympanic sinus of P. papillon sp. nov. is the most developed of any Perameles (most visible in paratype WAM M126), being wide with a high posterior wall.
The I5 is pointed and canine-like as in P. b. bougainville , P. b. notina , P. nasuta and P. pallescens . The canines are small with accessory cusps, in both males and females, as in P. b. bougainville , P. b. notina and P. eremiana (males of P. gunnii , P. nasuta and P. pallescens have an enlarged canine, and males of P. nasuta and P. pallescens generally lack accessory cusps). The canines are entirely bordered by the maxilla as in all Perameles , except P. eremiana (bordered by premaxilla anteriorly). The diastema between the canine and P1 is long (longer than P1) as in all Perameles except P. nasuta and P. pallescens . The P1 is as long as the P2 as in P. gunnii , P. nasuta and P. pallescens (shorter in P. b. bougainville , longer in P. eremiana and P. b. notina ), and the P2 is noticeably shorter than the P3, as in P. b. notina , P. nasuta and P. pallescens (subequal in length in all other Perameles ). The development of the lingual shelf of P3 is small as in P. b. bougainville and P. eremiana . The major cusp of P3 is similar in shape to all Perameles except P. gunnii (more conical). The upper molars of P. b. notina are completely worn down and the holotype lacks a dentary and therefore these are not used in the comparison following. On the M1, a short stylar crest is present anterior to the metastyle (best seen on juvenile WAM M572) with an associated StE, while StE is a conical cusp with no associated crest in P. b. bougainville (best seen in juvenile WAM M16086). StA is variable in size on M1–2. StB and StC are fused and indistinguishable as in P. b. bougainville and P. eremiana . The preparacrista reconnects with the postparacrista posteriorly on the M1, as in P. eremiana . The postprotocrista on M1 terminates posterior to the metacone as in P. b. bougainville and P. gunnii with no distinct posterior cingulum present (terminates more buccally in all other Perameles , as a posterior cingulum). On the M2, StB is conical as in all Perameles except P. gunnii (oval in shape). No stylar crest is present posterior to the conical StD as seen in P. gunnii and P. b. bougainville (small crest present in all other taxa). The postprotocrista ends posterior to the metacone as in all Perameles , except P. nasuta . StE is present as a distinct large cusp as in P. gunnii (it is reduced in all other species). On the M3, the postprotocrista ends posterior to the metacone as in all Perameles , except P. nasuta . The metaconule is small with only a small shelf separating it from the lingual flank of the metacone, as in P. b. bougainville and P. pallescens (a larger metaconule and shelf is present in all other taxa).
StE is absent as in all Perameles except P. gunnii . No anterior cingulum present as in all Perameles except for some specimens of P. pallescens and P. nasuta . On the M4, StB is variable, sometimes absent, sometimes present as a small cusp. This feature is variable across Perameles . The postparacrista is curved, forming a small centrocrista, as in all Perameles , except P. gunnii and P. b. bougainville , where it is straight. The postprotocrista ends anterior to the posterior end of the postparacrista as in all Perameles except P. nasuta . The metacone is small as in all Perameles except P. gunnii and P. nasuta , which have an enlarged metacone. StC/D is absent (sometimes present in P. gunnii and P. nasuta ).
The lower canine is small and has accessory cuspids as in all Perameles except in males of P. nasuta and P. pallescens , which have enlarged unicuspid canines. The diastema between the canine and p1 is shorter than the length of p1, as in P. pallescens (longer that p1 length in all other taxa). The p1 is as long as the p2 as in P. b. bougainville and P. eremiana (it is shorter in all other taxa). The p2 is longer than the p3 as in P. b. bougainville and P. eremiana (subequal in all other taxa). No cuspid is present on the hypoflexid of the lower molars as in all Perameles except P. nasuta and P. eremiana . On the m1, the paraconid-metaconid distance is equal to the metaconid-protoconid distance as in all Perameles except P. pallescens , which has a longer paraconid-metaconid distance. The posthypocristid is oblique to the tooth row and connects to the hypoconulid as in all Perameles (sometimes perpendicular in P. b. bougainville and connects to entoconid). On the m2, the posthypocristid is also oblique and connects to a large hypoconulid (this feature varies across Perameles ). On the m3, the hypoconid is buccal to the protoconid as in all Perameles except P. b. bougainville and some specimens of P. pallescens , which have the two cuspid level with one another. The posthypocristid connects to the entoconid as in all Perameles except some specimens of P. nasuta and P. pallescens , and the hypoconid is either absent or reduced as in P. b. bougainville and P. eremiana . On the m4, the talonid is reduced but not as reduced as in P. pallescens , P. b. bougainville and P. eremiana . The posthypocristid is perpendicular to the tooth row as in all Perameles except P. gunnii . The entoconid is large as in all Perameles , except P. b. bougainville and P. eremiana . The buccal shelf ends at the buccal side of the hypoconid as in P. b. bougainville and P. eremiana (buccally reduced in other taxa).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.