
Hyattella, Lendenfeld, 1888
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5361.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:EA087A82-BAC3-4316-9ABF-089421FD9051 |
|
DOI |
https://doi.org/10.5281/zenodo.10248064 |
|
persistent identifier |
https://treatment.plazi.org/id/03A72A36-D66D-FFCB-FF0C-C16EFD22FA38 |
|
treatment provided by |
Plazi |
|
scientific name |
Hyattella |
| status |
|
The Hyattella View in CoL issue
The genus Hyattella was established as a monotypic genus by Krantz (1962: 6), based on Hyattella epicrioides Krantz, 1962 as the type species, collected from Central Africa (Democratic Republic of the Congo), with the following brief diagnosis for the female: “Dorsal shield entire, without incisions; posterior part of shield with fifteen pairs of pectinate, distally expanded setae; vertical setae well developed; presternal shields present; pulvillar lobes rounded.” All of these characters are too general and fit several genera.
Krantz (1962) placed Hyattella in the subfamily Platyseiinae and the family Aceosejidae (now Blattisociidae ) along with Sejus C. L. Koch (now Cheiroseius Berlese ), Plesiosejus Evans & Hyatt , and Platyseius Berlese. Hyattella was placed in Platyseiinae primarily because of the elongated setae on the hypostome (which are actually normal) and palp trochanter, and the presence of the specialised strap-like macrosetae on the telotarsi II‒IV. Krantz considered the placement of his new genus within the subfamily as tentative and pointed out the absence of some typical features of Platyseiinae with respect to some structures of the leg ambulacrum and cheliceral dentition. Important apomorphic features of Platyseiinae include an acuminate median lobe of the pulvillus and elongate and acuminate paradactyli on the ambulacrum of the tarsi II‒IV (as an adaptation to movement in moist substrates), as well as some structures on the fixed cheliceral digit such as a deep subapical receptacle for the tip of the movable digit and a row of small to fine teeth confined to a ridge in the apical third of the masticatory surface ( Lindquist & Moraza 2016). Krantz (1962) also proposed that Hyattella represents an aberrant member of the aceosejine genus Lasioseius (based on the rounded pulvillar lobes, dorsal chaetotaxy, and cheliceral dentition) or a taxon with a position between Platyseiinae and Aceosejinae, and he systematically listed the genus Zerconopsis Hull as the most closely related group with respect to the modified strap-like macrosetae on the tarsi II‒IV.
Although the presence of tarsal macrosetae did not enter into the genus diagnosis, it was the main reason for Krantz to establish his new genus. However, Lindquist & Moraza (2016) evaluated the presence of these macrosetae as an expression of convergence rather than apomorphy. Currently, the genus is recognised by several authors as a synonym of the genus Lasioseius Berlese, 1916 ( Lindquist & Evans 1965; Walter & Lindquist 1997; Halliday et al. 1998). In addition, the generic name Hyattella Krantz, 1962 is a primary junior homonym and is occupied by Hyattella Lendenfeld, 1888: 233 (Porifera: Dictyoceratida : Spongiidae ), so this name is not available in the systematic nomenclature of Acari . Since Hyattella Krantz, 1962 has an available synonym ( Lasioseius Berlese, 1916 ), it is not necessary to establish the replacement name for this junior homonym (ICZN Article 60.1).
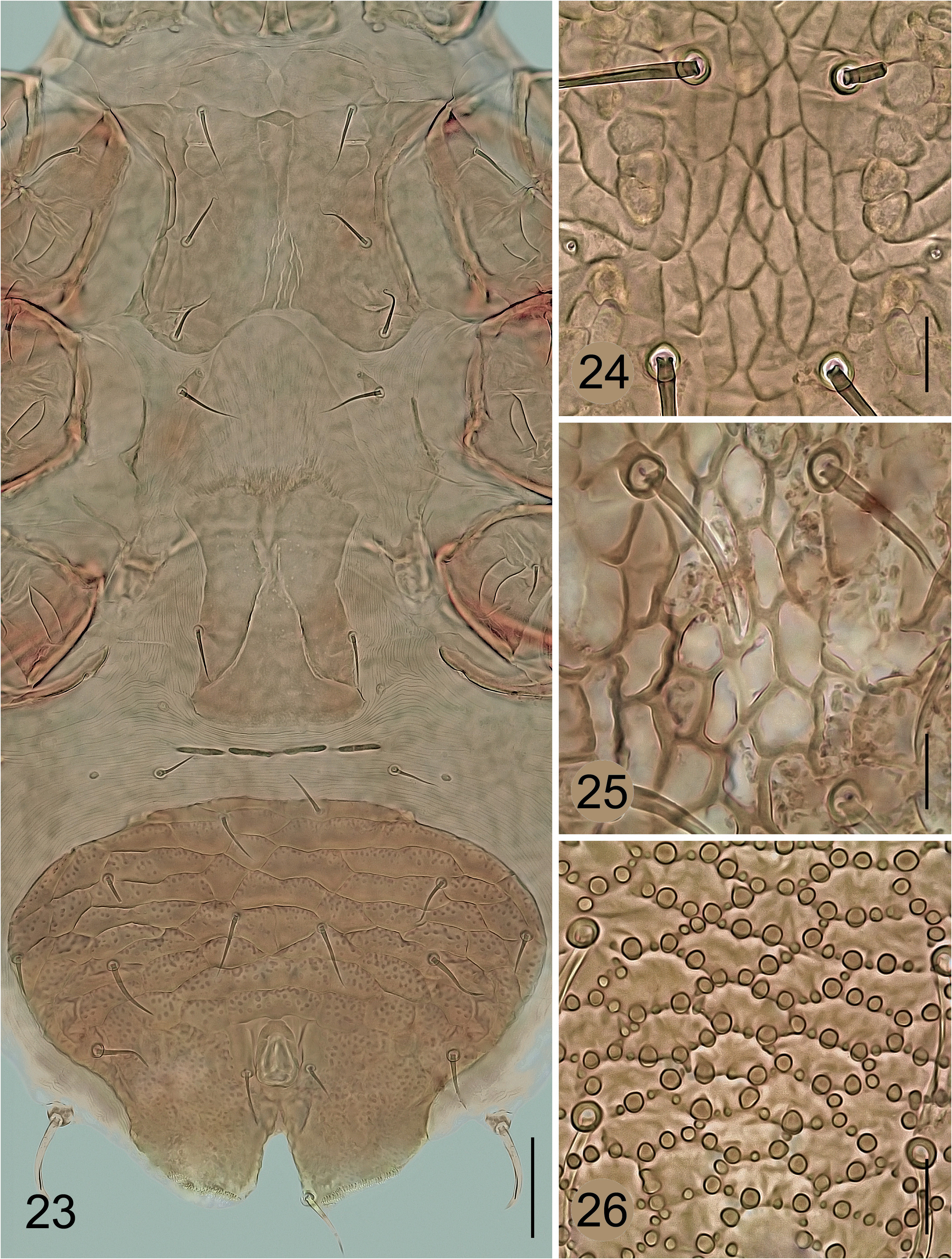
Since Lasioseius cheiroseioides (and its related species from the Palaearctic) also possesses specifically modified tarsal macrosetae on the legs II–IV as originally described in Hyattella epicrioides (actually Lasioseius epicrioides ), the type specimens of this type species of Hyattella were also examined to more accurately assess the mutual systematic affiliation of the two species. The examination and comparisons revealed some minor inconsistencies in the original description and illustrations of L. epicrioides , as well as significant morphological differences in some of the characters considered, despite the unusual presence of specialised tarsal macrosetae in both species ( Figs 26 View FIGURES 23–26 , 28 View FIGURES 27–31 , 32, 33 View FIGURES 32–33 ).
Krantz (1962) illustrated L. epicrioides with 38 pairs of setae on the dorsal shield and ten pairs of setae on the lateroventral soft cuticle, making no special comments on the number of setae in his description, and indicating and illustrating only five pairs of pre-anal setae on the ventrianal shield. In the holotype and the six paratypes available for my study (all females), I found 37 pairs of dorsal setae (all j / J, z / Z, and s / S setae, and r2‒r5), including the rather short and needle-like setae s1, s2, r2, and r4 (these setae are not modified and serrated like the other short but somewhat longer setae j2, z2, and J5); the ventrianal shield ( Fig. 33 View FIGURES 32–33 ) has six pairs of pre-anal setae (JV1‒JV4, ZV2, and ZV3) in addition to three circum-anal setae, and the soft cuticle has JV5, ZV4, ZV5, and 4‒5 pairs that could be considered both UR setae and R setae (one pair shown by Krantz on the soft cuticle at the level of the coxae III is actually on the dorsal shield). A similar species described from Angola by Aswegen & Loots (1969), L. dundoensis , appears to be identical to L. epicrioides , and its validity should be verified in a further study of the genus.
From my comparison of L. cheiroseioides and L. epicrioides , it appears that both species cannot be placed together in one species group in some further taxonomic concepts of the genus (if they are developed in the future). In contrast to L. cheiroseioides , the dorsal shield of H. epicrioides has a reticulate ornamentation with spherical tubercles ( Fig. 26 View FIGURES 23–26 ) and covers the entire dorsum and is unusually lateroventrally expanded to cover also the narrow lateral and posteroventral margins of the ventral idiosoma, the rows of marginal setae are incomplete, the presternal platelets are free from the sternal shield ( Fig. 32 View FIGURES 32–33 ), the metapodal region has only one pair of platelets, setae JV5 are short and needle-like, legs I are conspicuously long (including the tarsus) and significantly longer than legs IV, the chelicerae do not have a conspicuously elongated middle article, but the fixed digit is densely toothed along the entire masticatory surface, the epistome is tricuspid rather than triramous, the strap-like macrosetae of the tarsi II‒IV are straight (not curved and whip-like in the distal part), the tarsus III has only one macroseta (pd2 absent), and the macroseta on the tarsus IV is al2 instead of ad2.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.