
Athanas philippei, Anker & Ďuriš, 2022
|
publication ID |
https://doi.org/10.11646/zootaxa.5094.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:9655C635-FF92-4A87-AE47-2610CED08B7D |
|
DOI |
https://doi.org/10.5281/zenodo.6301624 |
|
persistent identifier |
https://treatment.plazi.org/id/03A387EF-2669-FFFA-E39B-FC4A79BAE9D7 |
|
treatment provided by |
Plazi |
|
scientific name |
Athanas philippei |
| status |
sp. nov. |
Athanas philippei n. sp.
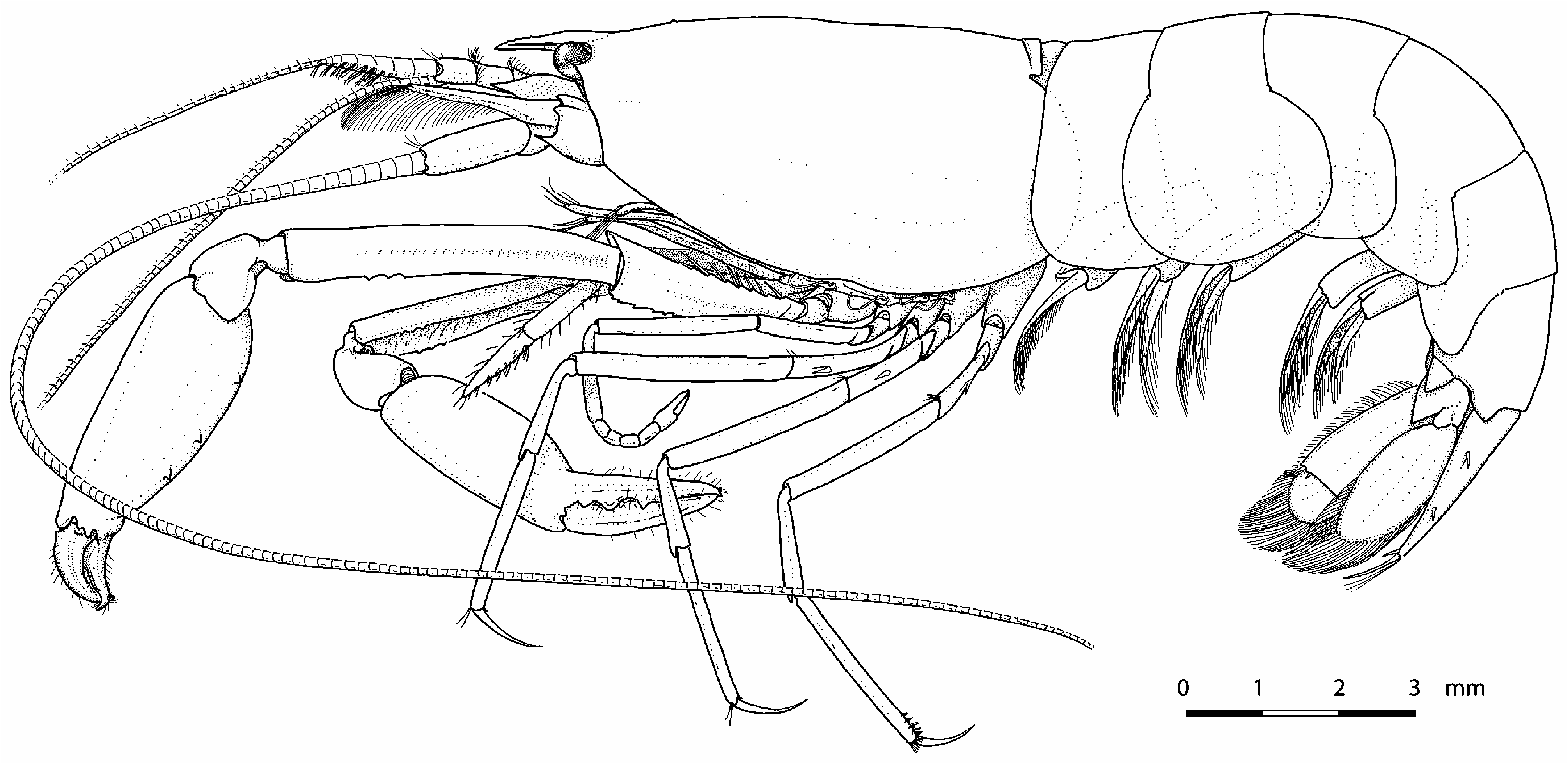
( Figs. 1–5 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 , 6A, B View FIGURE 6 )
Type material. Holotype: female (cl 6.9 mm), MNHN-IU-2018-1124, New Caledonia, north-eastern coast, Tiari , sta. KM608 , 20°15.4’S, 164°22.7’E, coastal platform composed of sand and fragmented coral rubble interspaced with shallow sandy pools along mangrove coast, depth less than 0.5 m, suction pump, in burrow, leg. A. Šobáňová & Z. Ďuriš, 23.09.2018. GoogleMaps
Paratypes: 1 female (cl 6.0 mm), MNHN-IU-2018-608, same collection details as for holotype; 1 female (cl 7.8 mm; freshly moulted), MNHN-IU-2018-1123, same collection details as for holotype; 1 female (cl 7.2 mm), MNHN-IU-2019-2986, New Caledonia, north-eastern coast, Saint-Mathieu , sta. KM606 , 20°22.8’S, 164°35’E, sandy coastal platform along mangrove coast, depth less than 0.5 m, suction pump, in burrow, leg. Z. Ďuriš, 01.11.2019 GoogleMaps ; 1 female (cl 5.2 mm), MNHN-IU-2018-1017, New Caledonia, north-eastern coast, Pouébo , sta. KM607 , 20°17.6’S, 164°29.1’E, shallow subtidal sand flat with nearby patches of seagrass and fragmented coral rubble, depth less than 0.5 m, suction pump, in burrow of Lysiosquillina maculata [host deposited under MNHN-IU-2018-1087], leg. A. Anker, 19.09.2018 GoogleMaps .
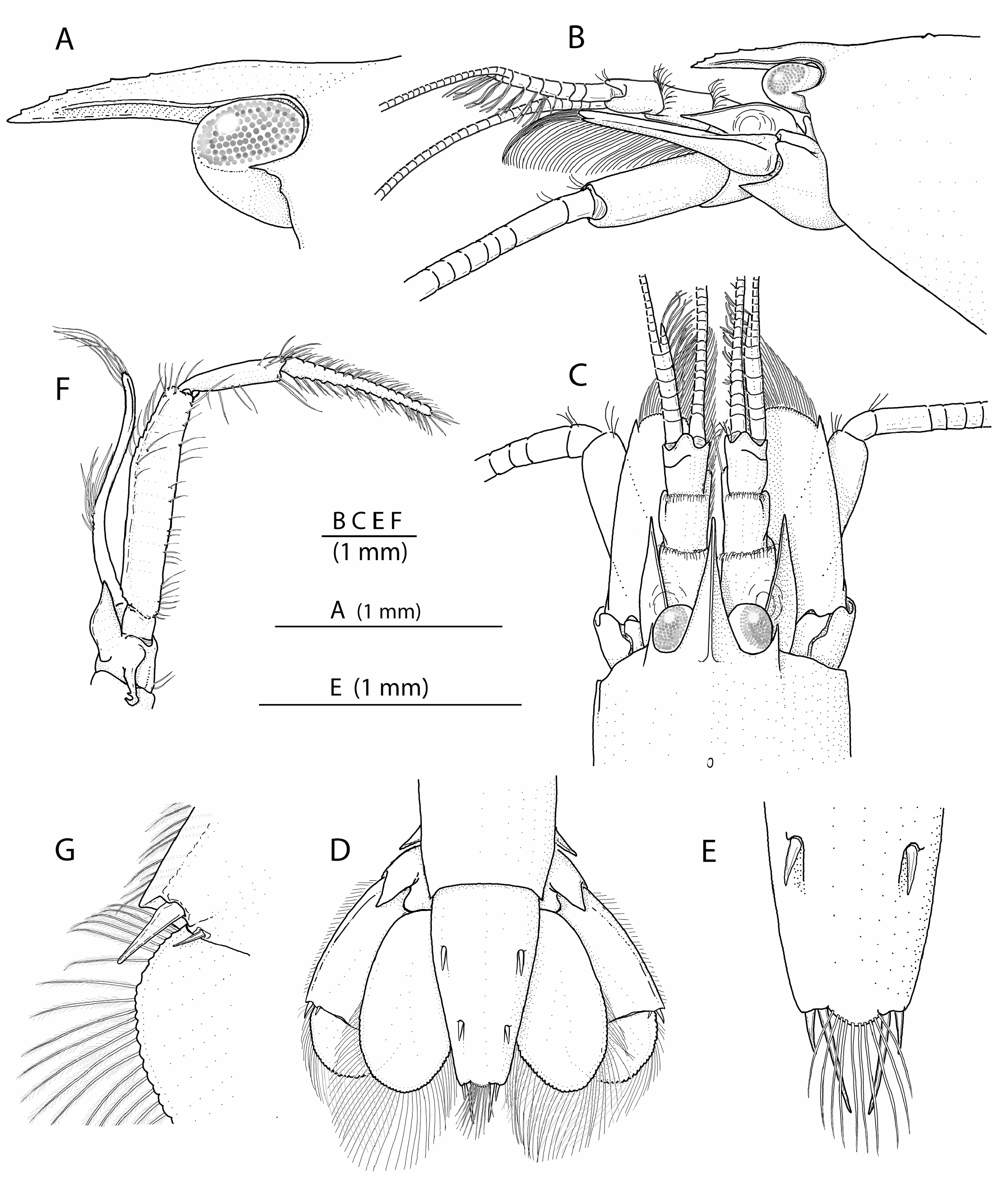
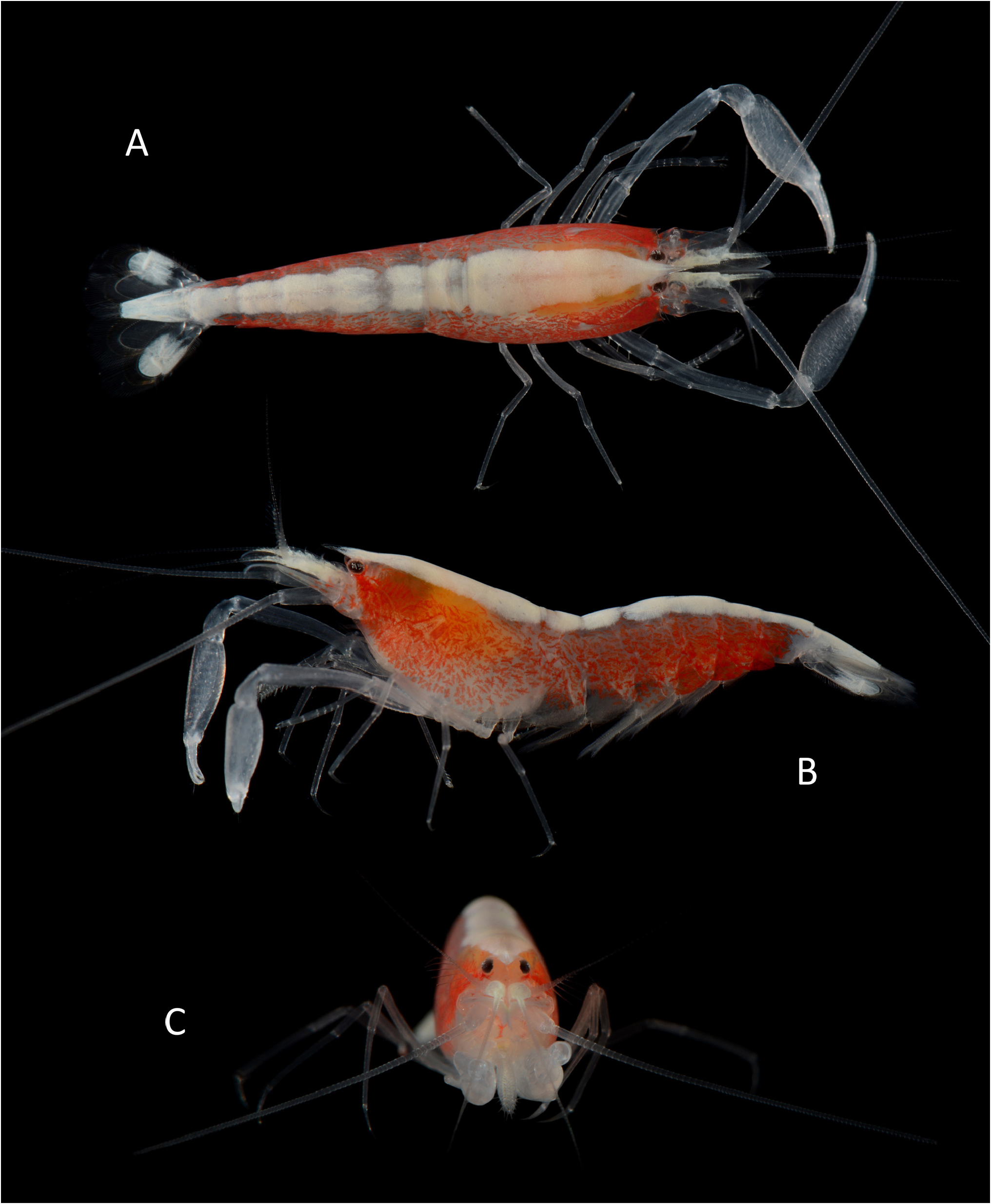
Description of holotype. Carapace ( Figs. 1 View FIGURE 1 , 2B, C View FIGURE 2 ) glabrous, with low median post-rostral tubercle and deep cardiac notch. Rostrum ( Fig. 2B, C View FIGURE 2 ) reaching to mid-length of second article of antennular peduncle, somewhat broadened at base, tapering distally; rostral carina distinct, flattening posteriorly, with 5 minute blunt teeth on anterior half. Extra-corneal teeth ( Fig. 2A–C View FIGURE 2 ) small, acute, laterally concealing basal-most portion of eye corneas. Infra-corneal teeth ( Fig. 2A–C View FIGURE 2 ) poorly developed as small bumps. Pterygostomial angle ( Fig. 2B View FIGURE 2 ) rounded, not or slightly produced anteriorly.
Pleon ( Fig. 1 View FIGURE 1 ) with first to fifth pleura rounded distoventrally; sixth pleonite with small subacute distal projection near telson base and subtriangular articulated plate. Telson ( Fig. 2D, E View FIGURE 2 ) subrectangular, slightly tapering distally, nearly twice as long as proximal width, with 2 pairs of stout cuspidate setae situated approximately at 0.3–0.4 (anterior) and 0.6–0.7 (posterior) of telson length; posterior margin slightly convex, with 2 pairs of spiniform setae, mesial more than twice as long as lateral.
Eyes ( Fig. 2A–C View FIGURE 2 ) large, largely exposed, with well-developed and normally pigmented corneas, latter occupying most of distal and distolateral area of eyestalk; anteromesial margin of eyestalks rounded, unarmed.
Antennule ( Fig. 2B, C View FIGURE 2 ) moderately stout; stylocerite relatively slender, with acute tip, reaching mid-length of second article; second and third article subequal in length; second article about 1.3 times as long as wide; lateral flagellum with fused portion composed of about 6 units, accessory ramus subdivided into 3 more or less distinct units, with several groups of aesthetascs.
Antenna ( Fig. 2B, C View FIGURE 2 ) with basicerite robust, armed with stout subacute distoventral tooth; scaphocerite broad, overreaching end of antennular peduncle; lateral margin straight; blade broadly convex anteriorly, slightly overreaching small distolateral tooth; carpocerite stout, overreaching antennular peduncle, but falling short of distolateral tooth of scaphocerite.
Mouthparts not dissected, typical for genus in external observation.Third maxilliped ( Fig. 2F View FIGURE 2 ) slender, pediform; coxa with distally subacute lateral plate; antepenultimate article about 5 times as long as greatest width; penultimate article slightly widening distally, about 4 times as long as distal width; ultimate article slender, tapering distally, with unarmed tip; exopod well developed, slightly overreaching antepenultimate article.
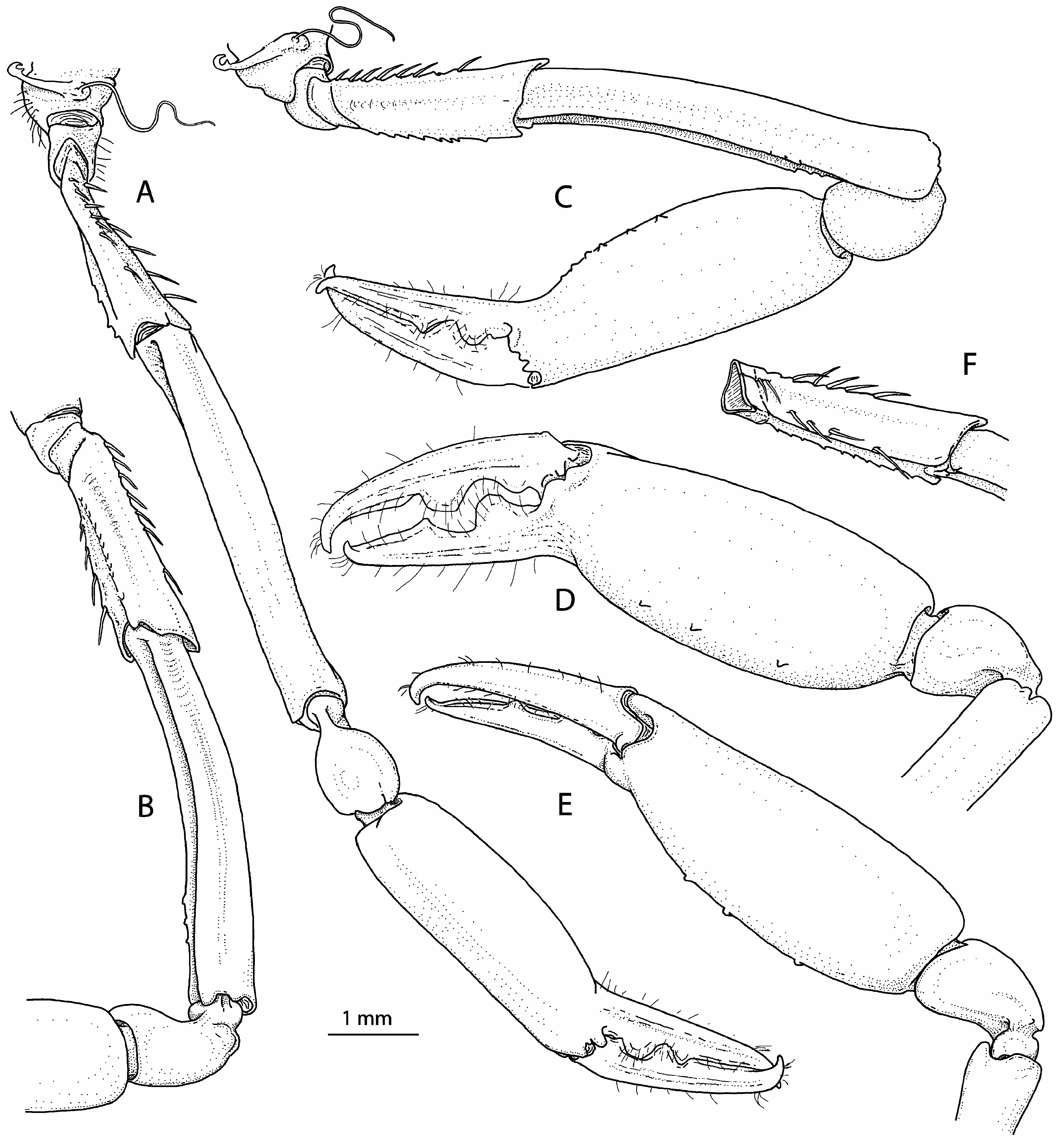
First pereiopods (= cheliped) subequal in size/length and somewhat asymmetrical in shape ( Fig. 3 View FIGURE 3 ), carried folded ventrally when at rest ( Fig. 5 View FIGURE 5 , 6A, B View FIGURE 6 ). Right (minor) cheliped ( Fig. 3A–C View FIGURE 3 ) moderately robust. Ischium somewhat flattened ventrally, with about 12 slender spiniform setae, most organised in 2 rows, 1 row along dorsal margin and 1 more irregular row on mesial surface, in addition to 1 spiniform seta situated on proximo-mesial surface; ventrolateral margin distinctly rugose. Merus relatively slender, almost twice as long as ischium, almost 8 times as long as wide at mid-length, not swollen, shallowly excavated ventrally; ventromesial and ventrolateral margins each with row of 3 poorly marked, low, blunt teeth subdistally. Carpus relatively short, cup-shaped, constricted proximally. Palm subcylindrical, slightly more than twice as long as maximal width, with row of 6 small tubercles on ventrolateral surface and shallow sinus near base of pollex. Fingers somewhat twisted away from main chela axis, subequal, about 0.6 length of palm; tips crossing when chela closed; cutting edge of pollex with 2 teeth, proximal tooth larger and more rounded, distal tooth smaller and more triangular, distal portion smooth; cutting edge of dactylus with 2 teeth, proximal tooth small and rounded, more distal tooth broader, situated at mid-length of cutting edge; all 4 teeth fitting into broad hiatuses on opposed margins.
Left (major) cheliped ( Fig. 3D–F View FIGURE 3 ) slightly more robust than right cheliped. Ischium, merus and carpus generally similar to those of right cheliped; chela slightly more robust and with somewhat different palm-finger ratio. Ischium with 11 spiniform setae, most arranged in 2 rows as on right cheliped, in addition to 2 spiniform setae on proximomesial surface. Merus with ventromesial margin bearing row of 3 low blunt teeth subdistally; ventrolateral margin unarmed subdistally, but with somewhat irregular margin at about distal third. Palm noticeably more swollen than that of right cheliped, with 3 tubercles disposed in row on ventrolateral surface and distinct sinus near base of pollex. Fingers proportionally somewhat longer compared to those of minor chela, about 0.7 length of palm, with dentition generally similar to that of right cheliped, but with more pronounced teeth.
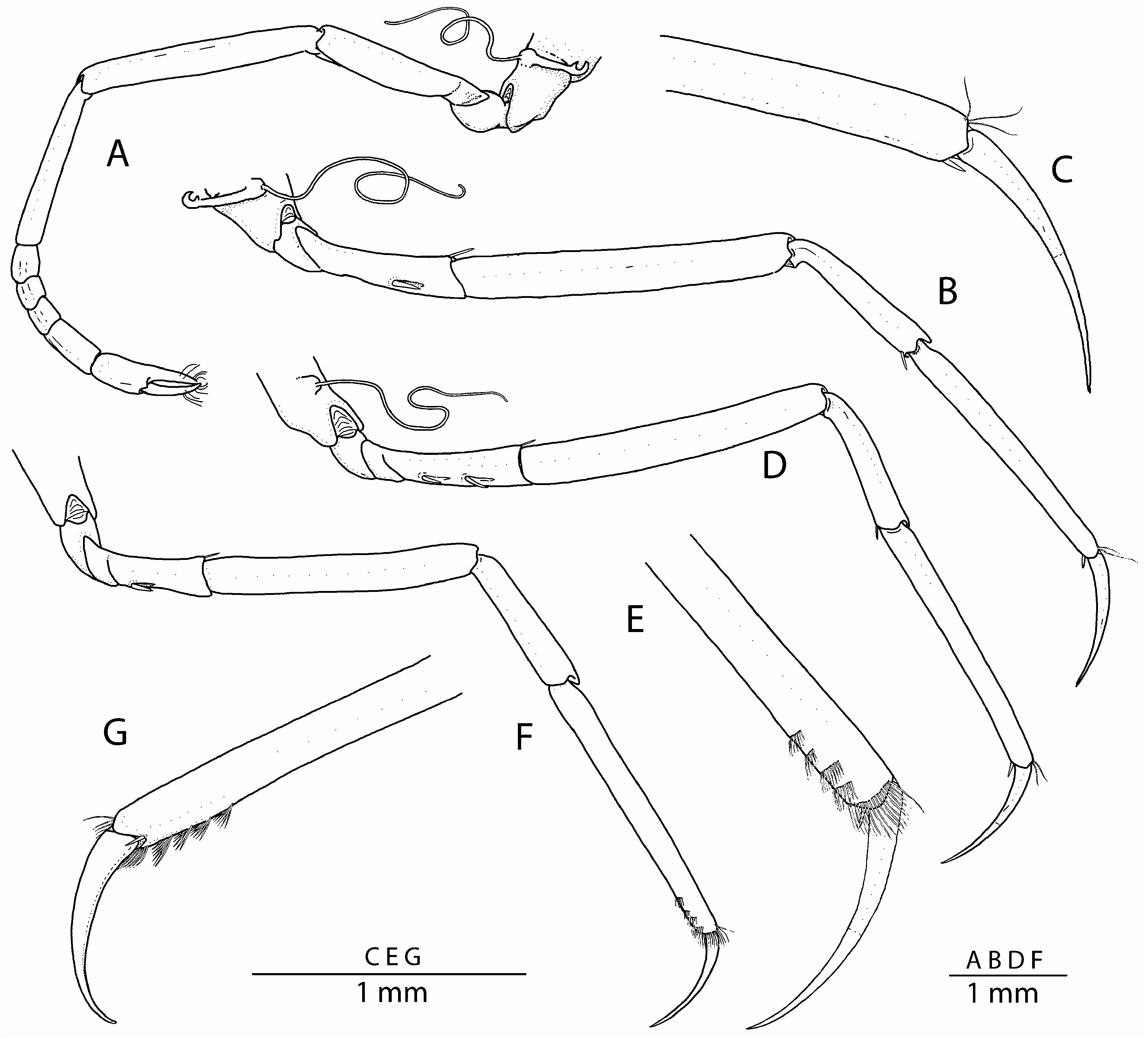
Second pereiopod ( Fig. 4A View FIGURE 4 ) slender; ischium about 5 times as long as wide; merus about 1.3 times as long as ischium; carpus slender, with five subdivisions, proximal slightly longer than sum of 4 others; chela simple, with fingers subequal to palm. Third pereiopod ( Fig. 4B, C View FIGURE 4 ) relatively slender; ischium about 5 times as long as wide, with 2 cuspidate setae on ventrolateral surface and 1 slender spiniform seta on distodorsal margin; merus somewhat slender, about twice as long as ischium and about 6.5 times as long as wide; carpus noticeably more slender than merus, about half as long as merus, with 1 slender spiniform seta on distoventral margin; propodus slender, unarmed ventrally, except for single small spiniform seta distally, near propodo-dactylar articulation; dactylus simple, slender, gently curved distally, almost half-length of propodus. Fourth pereiopod ( Fig. 4D View FIGURE 4 ) generally similar to third; ischium bearing 2 cuspidate setae on ventrolateral surface. Fifth pereiopod ( Fig. 4E, F View FIGURE 4 ) shortest and slenderest of walking legs; ischium with 1 cuspidate seta on ventrolateral surface and 1 slender spiniform seta on distodorsal margin; merus slightly more than twice as long as ischium; carpus about 0.6 length of merus; propodus slightly longer than merus, bearing 4 rows of serrulate grooming setae on distolateral surface and 1 pair of short spiniform setae distally, near propodo-dactylar articulation; dactylus slender, simple, curved, about 0.4 length of propodus.
Uropod ( Fig. 2D, G View FIGURE 2 ) with lateral lobe of protopod subacute; exopod with distolateral tooth and adjacent spiniform seta both rather small; lateral portion of diaeresis straight, with small, articulated, spiniform seta (referred to as spinule below) mesial to distolateral spiniform seta.
Gill-exopod formula: 5 pleurobranchs (P1–5); 0 podobranchs; 0 arthrobranchs; 4 strap-like epipods (Mxp3, P1–3); 4 setobranchs (P1–4); 3 exopods (Mxp1–3).
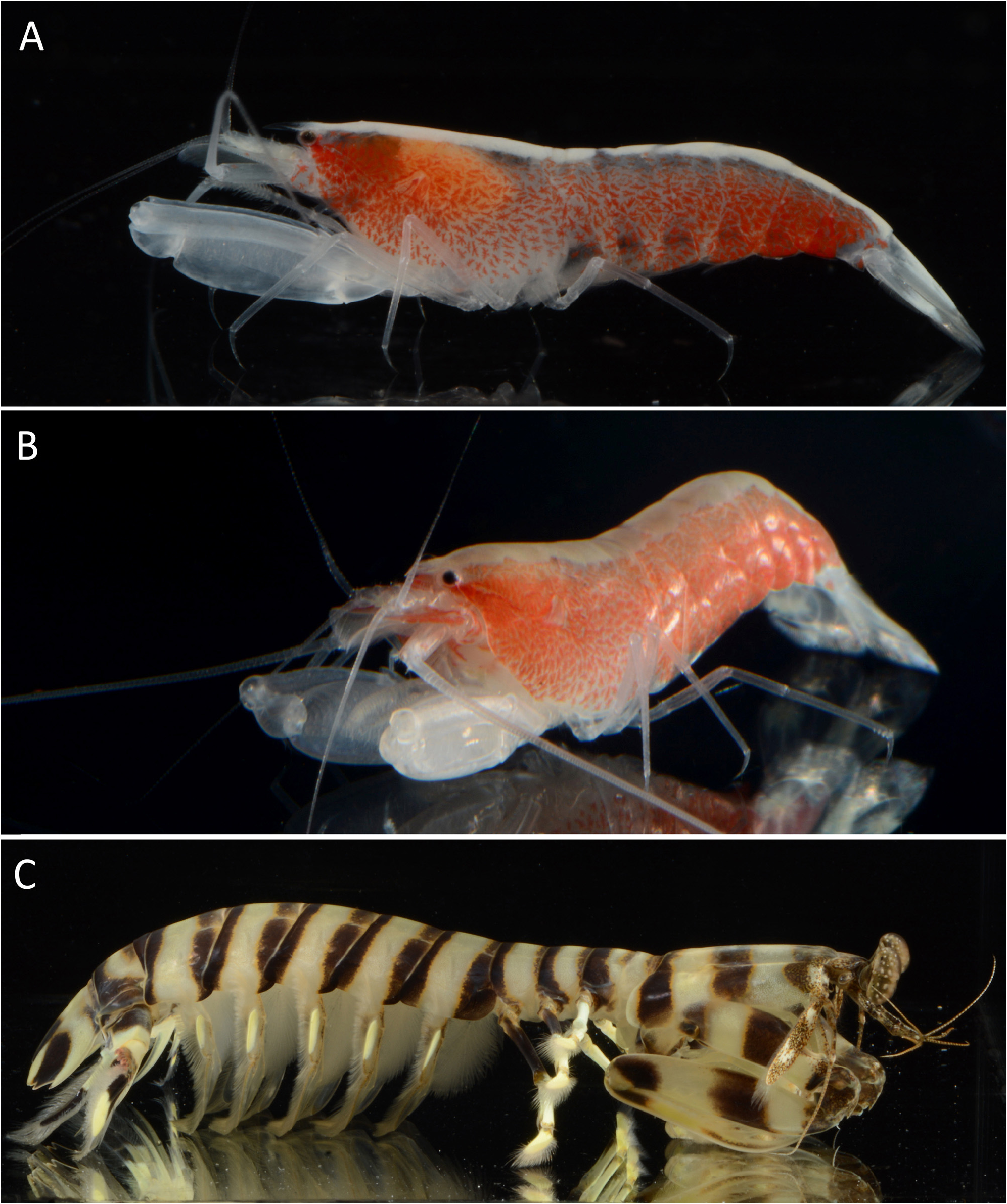
Colour pattern. Body generally semi-transparent; carapace and pleon flanks with more or less evenly distributed, irregular clusters of bright red chromatophores, some reminiscent of paired chromosomes of mammals, their density and colour intensity increasing towards posterior end of pleon; mid-dorsal area with broad, opaque, white or pale yellow band extending from tip of rostrum to telson, widest on carapace, tapering towards posterior pleonites; basal parts of antennae and antennules whitish with patches of red chromatophores; chelipeds hyaline white; pereiopods colourless, semi-transparent; telson entirely opaque white; uropod with transparent endopod and opaque white exopod ( Figs. 5 View FIGURE 5 , 6A, B View FIGURE 6 ).
Variation. The cl size range of the type series is 5.1–7.8 mm ( holotype: cl 6.9 mm). The rostrum typically reaches at least to the distal half of the second article of the antennular peduncle, as in the holotype, but falls short of it in some paratype specimens. The number of teeth on the rostral carina varies from 5 ( holotype) to 10 ( paratype MNHN-IU-2018-1123); in the latter case they extend almost to the base of the rostral carina ( Fig. 2I View FIGURE 2 ). The number of ventral tubercles on the chela palm varies from 1 (untypical condition, on both chelae of the paratype MNHN- IU-2019-2986) to 3 (minor) and 6 (major), as in the holotype and two paratypes (MNHN-IU-2018-608, MNHN- IU-2018-1087). The number of teeth on the ventrolateral margin of the cheliped merus varies from 3 ( holotype) to 6 ( paratype MNHN-IU-2018-608). The finger/palm length ratio of the minor chela is also variable to some degree, with the fingers being subequal to the palm in one of the paratypes (MNHN-IU-2018-608). The number of cuspidate setae on the ischia of the third to fifth pereiopods (P3–5) is relatively consistent: two on P3 and P4, and one on P5 (except on left P 3 in the paratype MNHN-IU-2019-1123, with a single spiniform seta). The small spinule on the uropodal exopod, which is present on both sides in the holotype, is present only on the right side in one paratype (MNHN-IU-2018-608), being replaced on the left side by a short, feebly plumose seta. This seta, instead of a spinule, is also present on both uropods in at least two other paratypes (MNHN-IU-2018-1123 and MNHN-IU- 2019-2986).
Etymology. The new species is named after our colleague, Prof. Philippe Bouchet (MNHN), a prominent malacologist and expert of marine biodiversity, and the main organiser of several “Our Planet Reviewed” expeditions, to which the authors were generously invited to participate.
Distribution. Presently known only from three localities on the north-eastern coast of New Caledonia, viz. Tiari ( type locality), Saint-Mathieu and Pouébo.
Ecology. Intertidal and shallow subtidal ( 0.5 m at low tide) sand flats adjacent to seagrass beds and coral rubble ( Fig. 7 View FIGURE 7 ); associated with burrows of the stomatopod Lysiosquillina maculata ; the host of the paratype MNHN-IU- 2018-1017 (deposited under MNHN-IU-2018-1087) had a total length of about 15 cm ( Fig. 6C View FIGURE 6 ).
Remarks. Athanas philippei n. sp. is morphologically closest to two other Indo-West Pacific species of Athanas , A. ahyongi (suspected to be associated with stomatopods) and A. manticolus (associated with Bigelowina phalangium , see below) ( Anker & Komai 2010; Ďuriš & Anker 2014).
The new species can be separated from A. ahyongi by the presence of a post-rostral tubercle on the carapace ( vs. absent in A. ahyongi ); the rostrum with a series of minute, spaced dorsal teeth ( vs. unarmed in A. ahyongi ); the rostrum reaching mid-length of the second article of the antennular peduncle ( vs. falling short of the distal margin of the first article in A. ahyongi ); the ultimate article of the third maxilliped very slender ( vs. much stouter in A. ahyongi ); the cheliped merus with few (typically three) low blunt teeth in subdistal position on both ventrolateral and ventromesial margins ( vs. armed with much stronger teeth along almost entire ventrolateral margin in A. ahyongi ); the ventral surface of the cheliped palm with one to six small but distinct tubercles ( vs. smooth in A. ahyongi ); and the third pereiopod propodus unarmed ventrally, except for a single distoventral spiniform seta (vs. with at least two additional spiniform setae on the ventral margin of the propodus in A. ahyongi ) (cf. Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ; Anker & Komai 2010). The colour patterns of A. philippei n. sp. ( Figs. 5 View FIGURE 5 , 6A, B View FIGURE 6 ) and A. ahyongi (cf. Anker & Komai 2010: fig. 6), although generally similar, are different in several important aspects. In the new species, the red chromatophores are present on most of the carapace and pleon flanks, whereas in A. ahyongi , they form a broad red lateral band, extending the full body length of the shrimp, with two colourless bands or areas, one between the red band and the white or pale-yellow mid-dorsal band, and another below the red band. Furthermore, in A. philippei n. sp., the uropodal exopod and endopod are opaque white and colourless, respectively, whereas in A. ahyongi , they are both red.
Athanas philippei n. sp. and A. manticolus share the presence of a minutely toothed rostrum, a feature also reported for the more distantly related A. dentirostris Anker, Jeng & Chan, 2001 , but generally highly unusual for alpheid shrimps ( Anker et al. 2001; Ďuriš & Anker 2014). However, the new species can be separated from A. manticolus by the noticeably longer rostrum, reaching to the middle of the second article of the antennular peduncle ( vs. reaching only to the distal margin of the first article in A. manticolus ); the extra-corneal teeth well-marked and acute ( vs. reduced to barely noticeable convexities in A. manticolus ); the cheliped ischium armed with several teeth on the ventrolateral margin ( vs. with one rather large triangular tooth in A. manticolus ); the cheliped merus with some low subdistal teeth on ventrolateral and ventromesial margins ( vs. with small teeth along entire ventrolateral margin in A. manticolus ); the cheliped carpus being much shorter, almost cup-shaped ( vs. elongate, more vaseshaped in A. manticolus ); the ventral surface of the cheliped palm with one to six relatively small tubercles ( vs. with one fairly large tubercle in A. manticolus ); the cheliped fingers armed with very stout teeth ( vs. with much weaker teeth in A. manticolus ); the noticeably slenderer third pereiopod, especially in the length-width ratio of the ischium and merus (for instance, merus about 6.5 times as long as wide in A. philippei n. sp. vs. about 4.5 times in A. manticolus ); and the ischia of the fourth and fifth pereiopods armed with two and one cuspidate seta(e), respectively ( vs. one and two, respectively, in A. manticolus ) ( Ďuriš & Anker 2014). The colour patterns of A. philippei n. sp. and A. manticolus are similar, especially in the red chromatophores clustering in short, randomly oriented lines on the sides of the body, the presence of a broad white to pale yellow mid-dorsal band and the mostly white tail fan. However, the broader chromosome-like clusters of A. philippei n. sp. are quite different from the narrower streaks of A. manticolus (cf. Figs. 5 View FIGURE 5 , 6A, B View FIGURE 6 ; Ďuriš & Anker, 2014: fig. 6A–C). In addition, the hosts of A. philippei n. sp. and A. manticolus are two quite different mantis shrimps. Lysiosquillina maculata (Lysiosquillidae) , the host of A. philippei n. sp. ( Fig. 6C View FIGURE 6 ), is a very large and powerful stomatopod popularly known as “tiger mantis shrimp” (maximal total length: 38 cm), which excavates large, spacious, U-shaped burrows ( Manning 1998; Ahyong 2001; Ahyong et al. 2017; A. Anker, pers. obs.). In contrast, Bigelowina phalangium (Nannosquillidae) , the host of A. manticolus ( Ďuriš & Anker 2014: fig. 6D), is a significantly smaller stomatopod (maximal total length: ca. 9.4 cm), which lives in much smaller U-shaped burrows ( Ahyong 2001).
Two other species of Athanas , viz. A. sydneyensis Anker & Ahyong, 2007 from south-eastern Australia and A. ivoiriensis Anker & Ahyong, 2007 from western Africa, share several morphological features with A. philippei n. sp., such as the general configuration of the frontal margin, the degree of symmetry and general shape of the chelipeds, and the proportions and armature of the third to fifth pereiopods ( Anker & Ahyong 2007). Athanas philippei n. sp. may be easily distinguished from both of them by the dorsally minutely toothed rostrum; the presence of a distinct post-rostral tubercle; and the reduced armature of the cheliped merus (which is armed with more or less strong teeth on the ventrolateral margin in the other two species). Athanas philippei n. sp. can be also distinguished specifically from A. sydneyensis by the presence of tubercles on the ventromesial surface of the palm ( vs. without tubercles in A. sydneyensis ); and from A. ivoiriensis by the ventrally unarmed carpus ( vs. with proximal teeth in A. ivoiriensis ) and finger cutting edges with a strong dentition ( vs. almost unarmed in A. ivoiriensis ) (cf. Anker & Ahyong 2007).
The presence of a small spinule mesial to the distolateral spiniform seta of the uropod in two specimens of A. philippei n. sp. ( Fig. 2D, H View FIGURE 2 ) is an interesting and possibly novel morphological feature within Athanas . This spinule is absent or at least has not been illustrated in A. ahyongi , A. manticolus , A. sydneyensis and A. ivoiriensis (cf. Anker & Ahyong 2007; Anker & Komai 2010; Ďuriš & Anker 2014), representing an additional distinguishing feature of the new species, albeit a somewhat inconsistent one, since it is absent on one or both uropods in some paratypes (see above).
None of the remaining species of Athanas appears to have close affinities to the present new species (e.g. Coutière 1903; De Man 1910; Holthuis 1951; Chace 1955; Banner & Banner 1960, 1966, 1973, 1978, 1983; Bruce 1990; Anker 2003; Anker et al. 2010; Marin 2017), including A. dentirostris , the only other species of the genus with a toothed rostrum, and A. squillophilus , a morphologically very distinctive species associated with stomatopods ( Anker et al. 2001; Hayashi 2002).
From an ecological point of view, A. philippei n. sp. is the fourth species of the genus Athanas directly encountered in a mantis shrimp burrow, suggesting a direct symbiotic association, the others being A. amazone , A. squillophilus and A. manticolus ( Froglia & Atkinson 1998; Hayashi 2002; Ďuriš & Anker 2014). Based on the presently available evidence, A. ahyongi also appears to be an inquiline shrimp, dwelling in stomatopod burrows, although no specimens have been collected so far together with a host ( Anker & Komai 2010). The hypothesis of the likely association between A. ahyongi and stomatopods is based on the collection data of the Nosy-Bé material (subtidal sand flat with rich infaunal community, including large stomatopods, AA, pers. obs.), the species’ general morphology, as well as its characteristic colour pattern, which is almost identical to the colour pattern of several shrimps, presumably Athanas sp. , photographed near entrances of stomatopod burrows (e.g. Kuiter & Debelius 2009: 159; Ryanskiy 2020: 29).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Caridea |
|
Family |
|
|
Genus |