
Arhochmus, Maslakova & Norenburg, 2008
|
publication ID |
https://doi.org/10.1080/00222930801995747 |
|
persistent identifier |
https://treatment.plazi.org/id/039CF65B-5440-FFA9-1947-197EFE80FE4B |
|
treatment provided by |
Felipe |
|
scientific name |
Arhochmus |
| status |
gen. nov. |
Arhochmus View in CoL gen. nov.
Type species
Prosorhochmus korotneffi Bürger, 1895 View in CoL .
Etymology
The name Arhochmus is formed by adding a negating ‘‘a’’ to a Greek RVXMOs (rhochmos)5cleft, runnel or gutter. It refers to lack of the prosorhochmid smile in this genus, in contrast to Prosorhochmus . The Greek has been Latinized by changing the terminal ‘‘os’’ to ‘‘us’’. The name is masculine in gender.
Diagnosis
Monostiliferous marine intertidal hoplonemerteans with rhynchocoel extending full body length, with wall composed of distinct outer circular and inner longitudinal muscle layers. Four simple eyes. Cephalic lobe is somewhat narrower than adjacent body region in actively moving worms. Proboscis small with a single pair of accessory stylet pouches and rounded basis of central stylet. Body-wall musculature well-developed with a delicate layer of diagonal muscles between outer circular and inner longitudinal muscle layers. Body-wall longitudinal muscle layer is not anteriorly divided. Frontal organ represented by a small epithelial pit. Cephalic glands extensive and contain at least three components: vacuolated basophilic mucus glands, finely granular acidophilic proteinaceous glands staining pink to dark red and coarsely granular proteinaceous glands staining yellow, golden brown or deep orange with Mallory trichrome or its modifications (orange-G glands). Cerebral sensory organs small with unforked canal, anterior or antero-lateral to brain, opening ventro-laterally into simple oblique cerebral organ furrows. Neurochord cells and neurochords absent. Lateral nerve cords without accessory nerve. Oesophagus lacking acidophilic glands. Caecum long, may be anteriorly bifid, with several lateral diverticula on each side. Anterior caecal diverticula reach posterior portion of brain. Blood system with three main longitudinal vessels, without transverse connectives. Mid-dorsal blood vessel with single vascular plug; cephalic blood loop planar (not recurved). Extracellular matrix of the so-called parenchyma scarce. Excretory system restricted to foregut region, with mononucleate terminal nephridial cells without distinct support bars, with thick-walled canals and up to six pairs of nephridiopores. Viviparous hermaphrodites.
Composition
Currently contains but a single species Arhochmus korotneffi ( Bürger, 1895) .
Geographic distribution
Currently only known from the Mediterranean coast of France (near Villefranchesur-Mer) and the Adriatic coast of Croatia (near Savudrija and Zambratija).
Arhochmus korotneffi ( Bürger, 1895) View in CoL comb. nov.
( Figures 1A, 1D, 1G View Figure 1 , 7C–D View Figure 7 , 8A–B View Figure 8 , 10 View Figure 10 ; Tables 1, 4) Prosorhochmus korotneffi View in CoL ( Bürger 1895, 1897 –1907, 1904; Oxner 1907a, 1907b; Friedrich 1955; Gibson 1982b, 1995)
Prosorhochmus claparedii ( Gibson and Moore 1985) View in CoL
Etymology
The species is named after Professor Korotneff, who first drew attention to the species and provided live and preserved specimens for description.
Type material
The type material was never designated and original specimens have almost certainly been lost. None of Bürger’s material, except two or three specimens, could be found, after exhaustive search efforts in likely European museums. In lieu of an available original type and with the purpose to clarify the taxonomic position of this species, we hereby designate a series of cross-sections of a gravid individual collected by SAM on Adriatic coast near Croatian coastal town Zambratija as a neotype. The neotype is deposited in the Smithsonian Institution’s National Museum of Natural History in Washington D.C., USA ( USNM 1020575 About USNM ) together with two additional sectioned ( USNM 1020573 About USNM , 1020574 About USNM ) and four unsectioned ( USNM 1025341 About USNM ) specimens collected from the same locality in Croatia .
Material examined
Arhochmus korotneffi ( Bürger, 1895) View in CoL . USNM 1020573-1020575, 1025341. Coll. SAM, Zambratija, Croatia.
Habitat and distribution
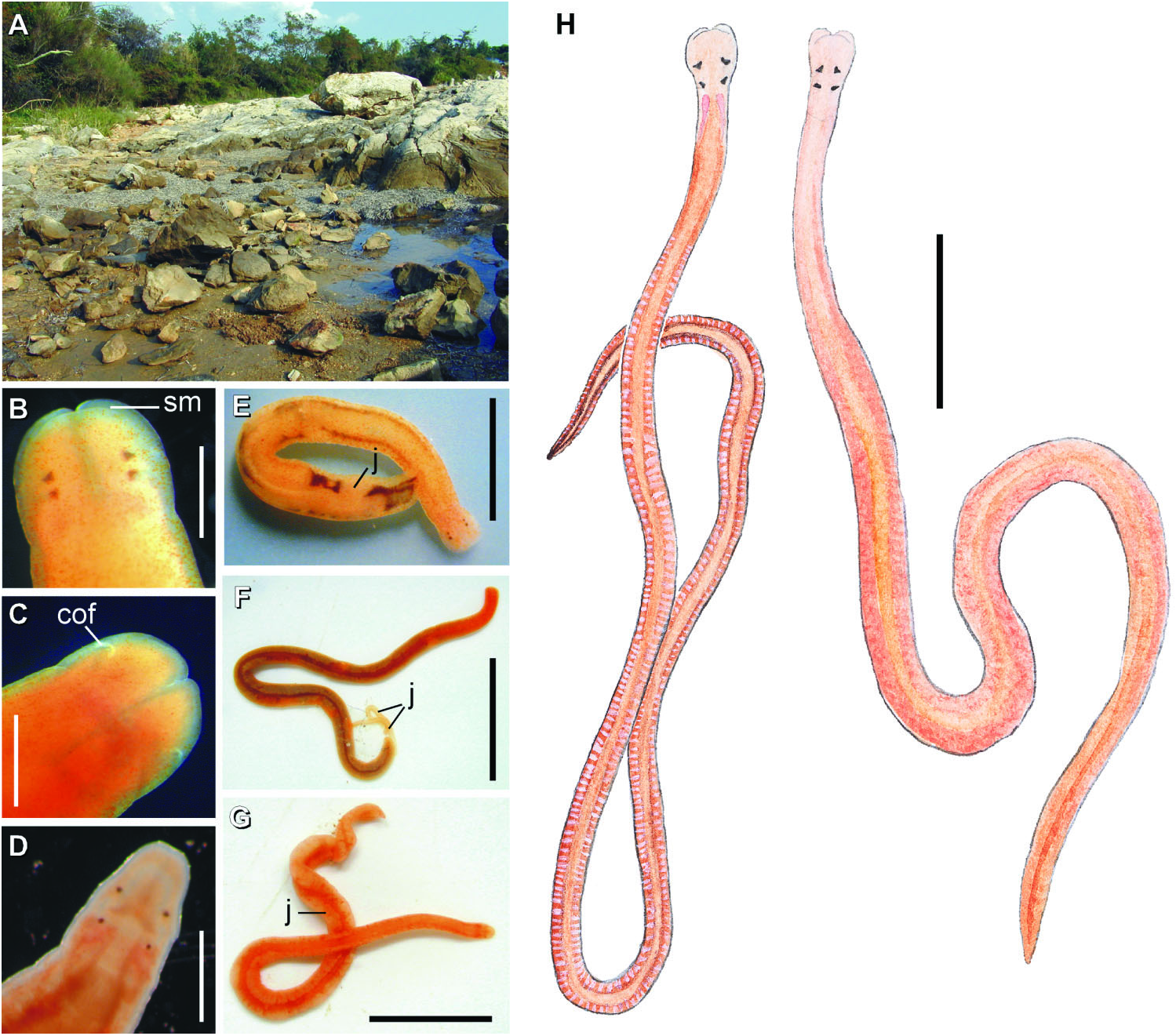
Intertidal to shallow sublittoral near Villefranche-sur-Mer, Provence, on the Mediterranean coast of France ( Bürger 1895). Additional specimens were collected by SAM from the coast of the Adriatic Sea near the Croatian coastal town Zambratija, in the north-western part of the Istrian Peninsula. Several individuals were found together crawling on the wet sand under the stones or on the lower surface of the stones just above the low water mark at low tide. The worms seem to prefer medium-size stones resting on the fairly coarse, somewhat muddy sand ( Figure 1A View Figure 1 ). Found in the same habitat as another viviparous hoplonemertean – Prosorhochmus chafarinensis .
Remarks
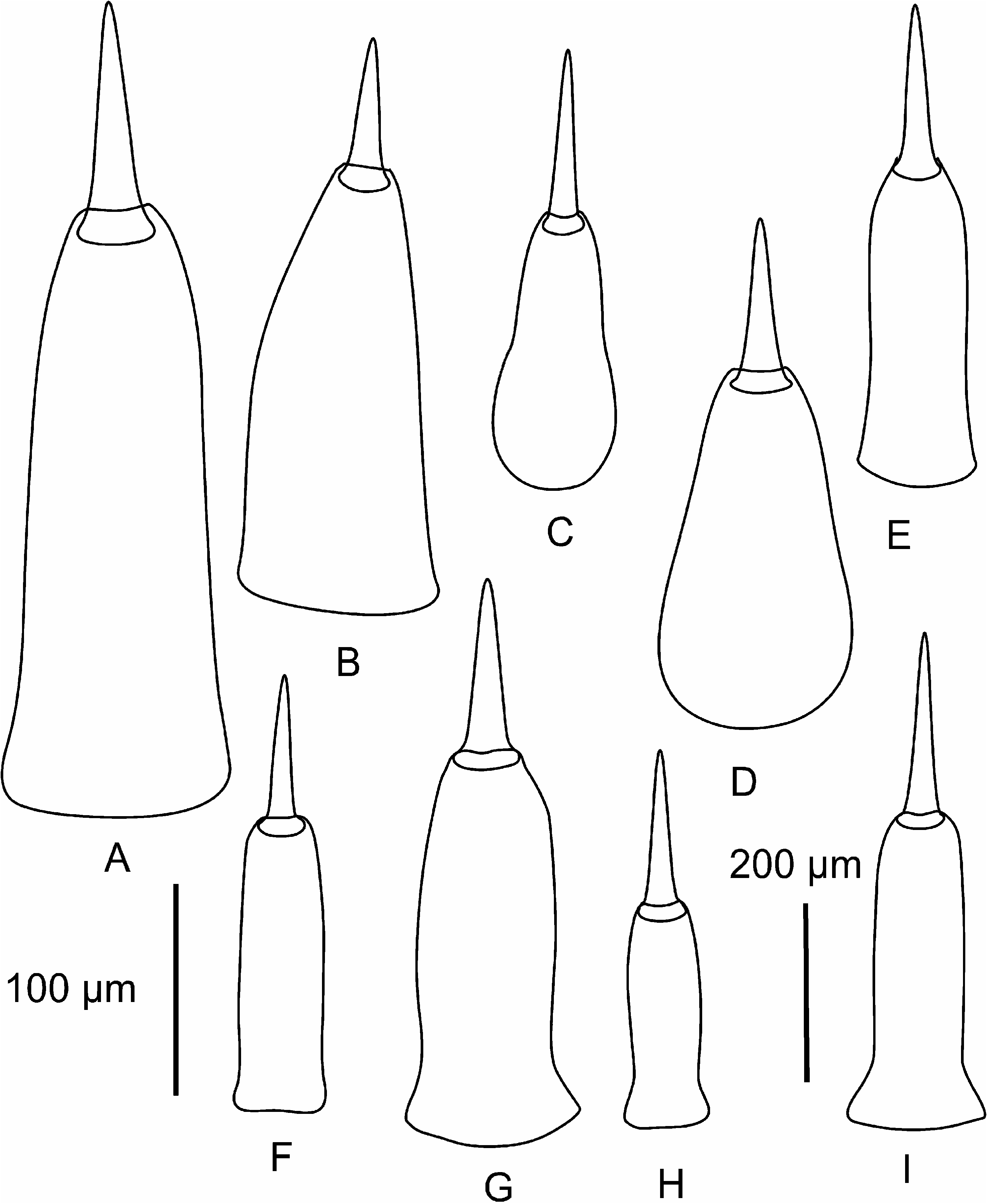
Arhochmus korotneffi ( Bürger, 1895) View in CoL is a viviparous, hermaphroditic intertidal species from the European coast of the Mediterranean Sea. Adults reach 60–70 mm in length and 1–1.5 mm in width. Body dorso-ventrally flattened, slender, tapering toward both ends. Colour in life is pinkish-orange, lateral margins of the body somewhat translucent, rhynchocoel appearing as a mid-dorsal stripe of deeper orange ( Figure 1G View Figure 1 ). Only four eyes have been observed in Croatian specimens ( Figure 1D View Figure 1 ), although Oxner (1907a) reported that there are occasionally five to seven eyes. Brooded young show through the body wall of the mature individual as large opaque, lighter-coloured areas between the intestinal diverticula ( Figure 1G View Figure 1 ). Proboscis with 12 proboscis nerves. Nephridia open via multiple distinct nephridiopores (up to six pairs). Central stylet (S) 90–110 Mm long, basis (B) pear-shaped, rounded, 115–170 Mm long; S/B ratio 0.65–0.83 (average 0.76) ( Figures 7C–D View Figure 7 ).
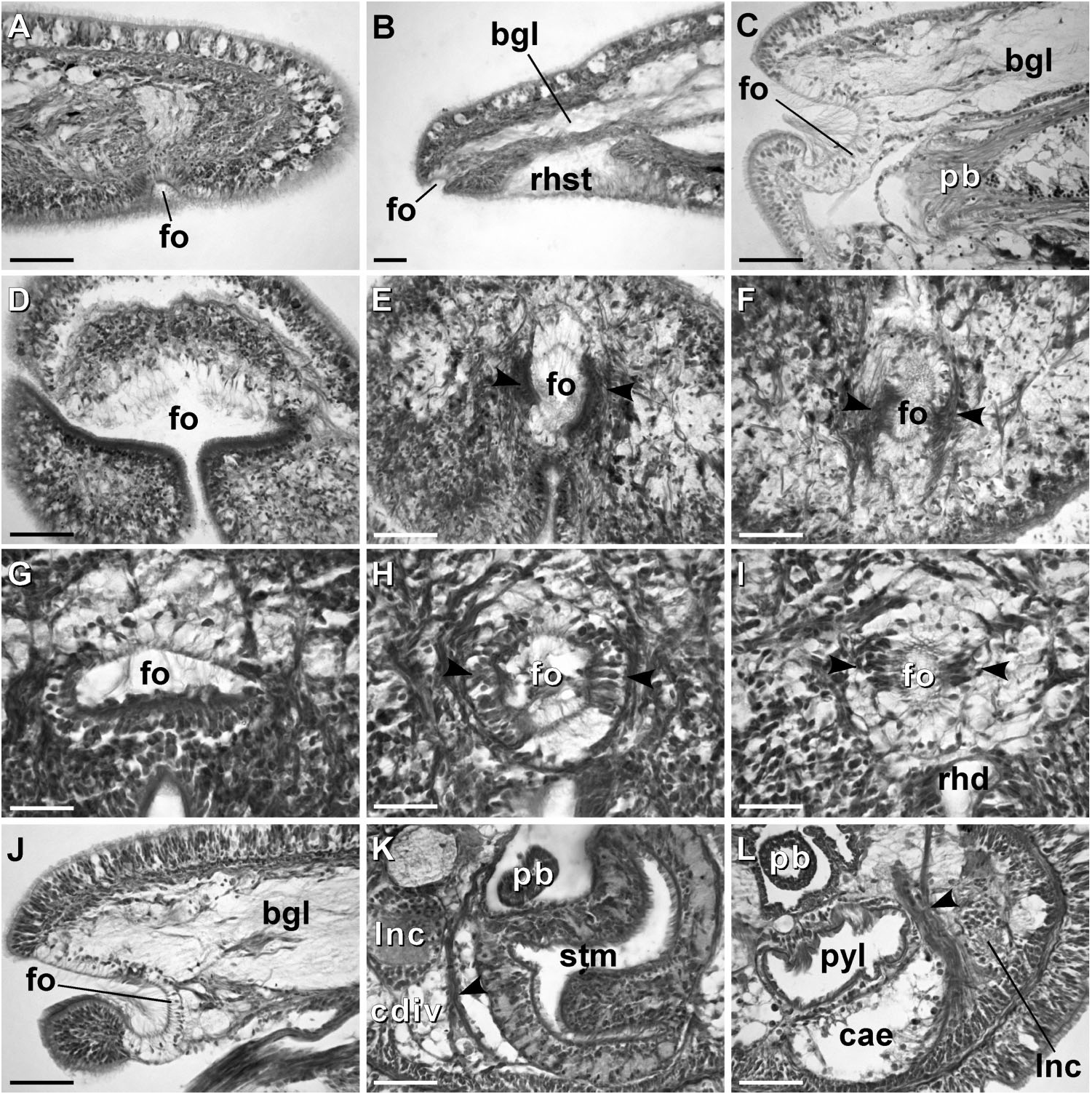
Despite the differences in head shape and stylet armature Gibson and Moore (1985) synonymized Prosorhochmus korotneffi Bürger, 1895 View in CoL with Prosorhochmus claparedii Keferstein, 1862 View in CoL , arguing that these characters may be intraspecifically variable. We have never observed these characters to vary intraspecifically in Prosorhochmus View in CoL or other nemerteans. Moreover, our specimens from Croatia fit Bürger’s (1895) description of Prosorhochmus korotneffi View in CoL exactly. Our observations of live specimens and serial histological sections suggest that P. korotneffi View in CoL is so morphologically different from P. claparedii View in CoL and other Prosorhochmus View in CoL that its inclusion in the genus is unwarranted. Unlike the true Prosorhochmus View in CoL , P. korotneffi View in CoL has a narrow head (vs. broad, bifid) without a prosorhochmid smile ( Figures 1D, 1G View Figure 1 , 10A–B View Figure 10 ), poorly developed frontal organ without lateral specialization of the epithelium of the frontal organ canal ( Figures 8A–B View Figure 8 ), several pairs of nephridiopores (vs. single pair) and, finally, a pear-shaped (vs. the typical truncated) central stylet basis of other Prosorhochmus View in CoL ( Figures 7C–D View Figure 7 ). Juveniles of P. korotneffi View in CoL are whitish in contrast to the yellowish-orange juveniles in P. claparedii View in CoL and Prosorhochmus chafarinensis View in CoL . The average sequence divergence between P. korotneffi View in CoL and other Prosorhochmus species is about twice as much as between the other species of Prosorhochmus View in CoL : 15.2% and 13.7% for 16S rDNA and COI, respectively ( Table 4). All of the above suggests that P. korotneffi View in CoL is not only distinct from P. claparedii View in CoL but that it is not a true Prosorhochmus View in CoL , despite its viviparity. Urged by our colleagues, we create a new monotypic genus Arhochmus View in CoL gen. nov. for Prosorhochmus korotneffi View in CoL .
Tetrastemma albidum ( Coe 1905a, 1905b)
Prosorhochmus albidus View in CoL ( Coe 1940, 1944; Friedrich 1955; Corrêa 1964; Gibson 1982b, 1995; Gibson and Moore 1985)
Etymology
The specific epithet refers to the milky-white colour of live specimens.
Type material
Type material had not been designated. One specimen found in Coe’s collection and identified as Prosorhochmus albidus by Coe was sectioned by Frank Crandall ( Washington, D.C.) and is currently deposited with the Smithsonian Institution’s National Museum of Natural History, Washington, D.C., USA ( USNM 126765 About USNM ) .
Material examined
‘‘ Prosorhochmus ’’ albidus – series of transverse sections, USNM 126765; additional material: numerous live specimens coll. by SAM and JLN at Bird Rock, near La Jolla, CA, USA ( Tetrastemma sp. 644, deposited at the National Museum of Natural History in Washington D.C., USA).
Habitat and distribution
Roots of sea grass, kelp holdfasts and coralline algae in the intertidal zone between Monterey Bay, California, USA and Ensenada, Mexico.
Remarks
Tetrastemma albidum , currently referred to as Prosorhochmus albidus (Coe, 1905) View in CoL , is a small milky-white or cream-coloured four-eyed hoplonemertean reaching maximum length of 25 mm, found in the intertidal zone between Monterey Bay, California and Ensenada, Mexico. The species was originally described as Tetrastemma albidum ( Coe 1905a) and subsequently transferred to Prosorhochmus View in CoL because it appeared to be occasionally viviparous, i.e. while most specimens were oviparous, some retained fertilized eggs within the ovaries until juveniles were formed ( Coe 1940). Gibson and Moore (1985) commented that P. albidus View in CoL was insufficiently described to determine its taxonomic status, however, Gibson (1995) included P. albidus View in CoL as a valid species. Several characters described and illustrated by Coe (1905a, p. 294. pl. 17, figs 104–105, pl. 22, figs. 145–149; 1940, p. 294, pl 25, fig. 20) make prosorhochmid affinity of this species doubtful: the head is narrow, not anteriorly bifid and lacks a prosorhochmid smile, basis of the central stylet is rounded (compared to truncated bases of all other Prosorhochmus View in CoL ) and, finally, cerebral organs are large and extend ventro-laterally to the posterior portion of brain (compared to small cerebral organs, located anteriorly to brain in true Prosorhochmus View in CoL ).
We collected numerous specimens of small four-eyed cream-coloured hoplonemerteans from the roots of sea grass, kelp holdfasts and coralline algae in January 2002 at Bird Rock, La Jolla, CA, which appear to fit Coe’s description (1905a). Specimens collected by us were 10–15 mm long, whitish or cream-coloured, with a typical tetrastemma-like narrow head without anterior notch or a prosorhochmid smile; and base of the central stylet was rounded. These specimens also resembled Coe’s description (1905a, 1940) by having a colour pattern on the head formed by diffuse brownish pigment spots. This pattern was quite variable: two pigment spots covering the anterior eyes, sometimes connected; four pigment spots – one over each eye, anterior spots connected, while posterior not; posterior spots connected while anterior not; anterior pair of eyes entirely covered with pigment in addition to diffuse pigment lines connecting anterior and posterior eyes of each side. Our specimens fixed for histology are deposited at the Smithsonian Institution’s National Museum of Natural History ( Tetrastemma sp. 644). If it is Coe’s Prosorhochmus albidus View in CoL , it definitely does not belong to Prosorhochmus View in CoL , based on the characters of external appearance and stylet apparatus.
Another line of evidence supporting non-prosorhochmid affinity of Prosorhochmus albidus comes from investigation of material that was found in Coe’s collection and identified as P. albidus by Coe. This material was sectioned by Frank Crandall and is currently deposited at the Smithsonian Institution’s National Museum of Natural History (USNM 126765). The lack of well-developed frontal organ, lack of lateral differentiation of frontal organ epithelium and lack of prosorhochmid smile; poorly developed basophilic mucus cephalic glands; lack of orange-G glands; and, finally, large cerebral organs situated postero-lateral to brain indicate that this species does not belong to Prosorhochmus or the family Prosorhochmidae .
We are urged by colleagues not to return P. albidus to its original home, the potpourri that is Tetrastemma . However, until there is an acceptable revision and diagnosis of Tetrastemma , it cannot be determined that T. albidum would not properly be associated with the designated type species of the genus. Until such time, there is no increased functional benefit to erecting what would be a monotypic genus of convenience – we know as little about systematic affiliation either way. We believe it is best to re-establish the original combination Tetrastemma albidum for P. albidus .

| SAM |
South African Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
Arhochmus
| Maslakova, Svetlana A. & Norenburg, Jon L. 2008 |
Arhochmus korotneffi ( Bürger, 1895 )
| Maslakova & Norenburg 2008 |
Arhochmus korotneffi ( Bürger, 1895 )
| Maslakova & Norenburg 2008 |
Arhochmus korotneffi ( Bürger, 1895 )
| Maslakova & Norenburg 2008 |
Arhochmus
| Maslakova & Norenburg 2008 |
Tetrastemma albidum
| Coe 1905 |
Tetrastemma albidum
| Coe 1905 |
Prosorhochmus korotneffi Bürger, 1895
| Burger 1895 |
Prosorhochmus korotneffi
| Burger 1895 |
Prosorhochmus korotneffi Bürger, 1895
| Burger 1895 |
Prosorhochmus korotneffi
| Burger 1895 |
P. korotneffi
| Burger 1895 |
P. korotneffi
| Burger 1895 |
P. korotneffi
| Burger 1895 |
P. korotneffi
| Burger 1895 |
P. korotneffi
| Burger 1895 |
Prosorhochmus korotneffi
| Burger 1895 |
Prosorhochmus claparedii
| Keferstein 1862 |
P. claparedii
| Keferstein 1862 |
P. claparedii
| Keferstein 1862 |
P. claparedii
| Keferstein 1862 |