
Kalanchoe, , Descoings, 2003
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.482.2.1 |
|
persistent identifier |
https://treatment.plazi.org/id/039C87D7-FF8D-CD64-FF32-D954FD84FBB3 |
|
treatment provided by |
Marcus |
|
scientific name |
Kalanchoe |
| status |
|
2. Kalanchoe View in CoL × rechingeri Raym. -Hamet ex Rauh & Hebding in Rauh (1995a: 17), pro sp.
Type:— MADAGASCAR. Toliara province (now the Anosy region), Préfecture de Fort-Dauphin [now Tôlañaro, alternatively spelled ‘Tolagnaro’], cultivated at [the private botanical garden of Julien Marnier-Lapostolle] “Les Cèdres”, [plant] “windend und kletternd” [“winding and climbing”], collected on 05 December 1993 [probably at “Les Cèdres”], W. Rauh s.n. [old HEID accession number 74214] (holotype, HEID HEID702311 View Materials [Image available at https://gartenbank.cos.uni-heidelberg.de/public/gb.php?sid=SID&gn=7 02311]) .
Note on the type:—The protologue of the name K. rechingeri contains a typographical error as the following is stated: “ Lectotype: B. G. H. 74213 dans l’herbier de l’Inst. Syst. Bot. de l’Univ. de Heidelberg ( HEID).” Rather, the material that Rauh & Hebding in Rauh (1995a: 18) must have intended to cite under K. rechingeri is B. G. H. 74214, i.e., the final ‘ 4 ’ in this number was incorrectly transcribed as ‘ 3 ’. This is the only specimen cited by Rauh & Hebding in Rauh (1995a: 18), and we here correct “ lectotype ” to “ holotype ”.
A specimen deposited at Herb. P, [P-P00431390; http://coldb.mnhn.fr/catalognumber/mnhn/p/p00431390], bears a specimen label that claims it to be an “isotype”, i.e., a duplicate of the holotype, of the name K. rechingeri . However, there is no evidence that this specimen, deposited under L. Allorge 1020, was collected at the same place and at the same time as the holotype. This specimen, collected on an unknown date, was donated to Herb . P by W. [Werner] Rauh, without a Rauh number, on 20 February 1995 , while the holotype is dated 05 December 1993.
Description:—see Rauh (1995a: 17), with the following amendments: margins only dentate apically, entire elsewhere, few-toothed, teeth subducted by pale organogenic and/or embryogenic pedestals, and, often bulbils proper, or abscission scars after bulbil detachment; bulbil production constitutive.
Additional material examined:—W 1954-0007470 [https://w.jacq.org/W19540007470]; P-P00444058 to P- P00444061 are possibly also representative of K. × rechingeri .
Parentage:— K. costantinii × K. tubiflora .
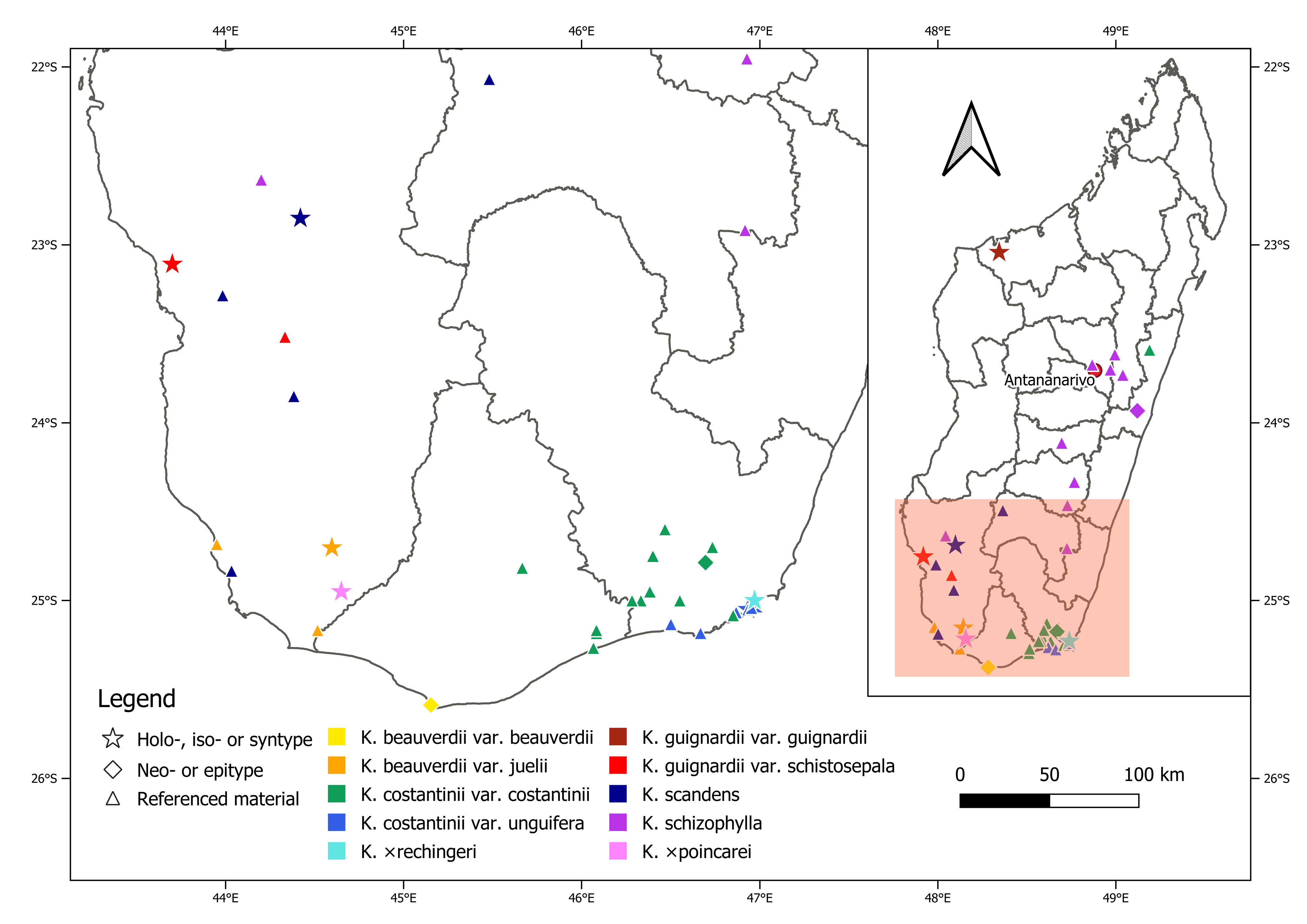
Distribution:—Around Tôlañaro, southern Androy region, southeastern Madagascar ( Fig. 7 View FIGURE 7 ).
Discussion:—Though originally described as a species, we agree with later works that treated K. × rechingeri ( Fig. 5C–F View FIGURE 5 ) as being of hybrid origin instead, particularly of the formula K. beauverdii × K. tubiflora in which K. beauverdii represents a broad concept ( Boiteau & Allorge-Boiteau 1995: 87, Descoings 2005a: 3). Curiously, despite proposing it to be a hybrid, Boiteau & Allorge-Boiteau (1995: 86) included it in their circumscription of K. beauverdii .
Under the taxonomy followed in this paper, of the species recognised in K. ser. Vilana, K. costantinii resolves as one of the parents of K. × rechingeri , based on the following traits:
1. The calyx of K. × rechingeri is fused for over half its length, i.e., the free sepal segments are shorter than the calyx tube, as is the case in K. costantinii . In contrast, in both K. tubiflora and all varieties of K. beauverdii the free sepal segments are longer than the calyx tube. The calyx tube of K. × rechingeri is also closer in shape to K. costantinii than to K. beauverdii .
2. The free corolla segments of both K. tubiflora and K. beauverdii are more obovate and broader towards the apex than those of either K. costantinii or K. × rechingeri .
3. In the vicinity of Tôlañaro [Tolagnaro], the distribution range of K. × rechingeri is sympatric with those of K. marnieriana , K. sanctula , K. tubiflora , and K. costantinii , as well as with some forms of K. rosei (see Humbert 1933: 166), but not with K. beauverdii ( Fig. 7 View FIGURE 7 ).
We agree with previous works that the other parent of K. × rechingeri is K. tubiflora , based on the following arguments:
1. All known K. tubiflora hybrids bear darkly maculate petioles, a trait overwise not observed among other representatives of K. sect. Invasores, suggesting it to be a dominant trait. This colour pattern, absent in sympatric forms of K. rosei and in all species recognised in K. ser. Vilana, is very distinct in K. × rechingeri ( Fig. 5D View FIGURE 5 ).
2. Kalanchoe × rechingeri , K. costantinii , and K. tubiflora all produce bulbils constitutively, a trait which we believe to be recessive or co-dominant based on its presence in other known hybrids of which K. tubiflora is one parent, i.e., K. × houghtonii, K. × richaudii, and “ K. Rauhii ”. In contrast, sympatric forms of K. rosei are induced-bulbiferous, and in a hybrid with K. costantinii , should produce an induced, or at most a semi-constitutively, bulbiferous hybrid, which is not the case for K. × rechingeri . Moreover, the leaf apices of K. × rechingeri bear bulbil pedestals ( Fig. 5D View FIGURE 5 ), which are present in K. tubiflora and K. sanctula , but not in K. costantinii or K. rosei .
3. The base of the calyx in K. × rechingeri , as well as in K. costantinii and especially in K. tubiflora is round ( Figs 2A–B View FIGURE 2 , 5E View FIGURE 5 ), while it tends to be less so to occasionally flattened in sympatric forms of K. rosei . Furthermore, those forms of K. rosei have a papery calyx texture, unlike the succulent calyx of K. × rechingeri , K. costantinii , and K. tubiflora .
4. The flowers of K. × rechingeri are large, rather inflated, and strongly spreading ( Fig. 5F View FIGURE 5 ), traits notable in all the known hybrids of K. tubiflora , i.e., K. × houghtonii, K. × richaudii, and “ K. Rauhii ” [a ‘name’ applied in horticulture]. In contrast, sympatric forms of K. rosei show rather elongated, subcylindrical calyces and corollae, the latter of which are also less spreading and less constricted at the base of the petals. Species of K. ser. Vilana have almost funnelform corolla tubes that are not much constricted at the base of the petals ( Fig. 6 View FIGURE 6 ), while K. tubiflora shows strongly spreading petals, a significant constriction at the base, and a strongly inflated upper corolla tube. Thus, K. × rechingeri shows an intermediate state in floral traits between K. costantinii and K. tubiflora , but not with sympatric forms of K. rosei .
Finally, the very narrowly-oblong, sublinear leaf morphology of K. × rechingeri is intermediate between the linear leaves of K. tubiflora and the oblong-ovate leaves of K. costantinii . This is not the case compared to other sympatric species, such as K. marnieriana or K. sanctula . The analysis above suggests that K. × rechingeri is a natural hybrid with the formula K. costantinii × K. tubiflora .



| W |
Naturhistorisches Museum Wien |
| HEID |
University of Heidelberg |
| B |
Botanischer Garten und Botanisches Museum Berlin-Dahlem, Zentraleinrichtung der Freien Universitaet |
| G |
Conservatoire et Jardin botaniques de la Ville de Genève |
| H |
University of Helsinki |
| A |
Harvard University - Arnold Arboretum |
| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
| L |
Nationaal Herbarium Nederland, Leiden University branch |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.