
Bos gaurus Hamilton-Smith, 1827
|
publication ID |
https://doi.org/ 10.1093/mspecies/sey004 |
|
publication LSID |
lsid:zoobank.org:pub:0F35A937-881D-459D-A9E5-6EBF32278A01 |
|
DOI |
https://doi.org/10.5281/zenodo.4592241 |
|
persistent identifier |
https://treatment.plazi.org/id/039587B7-1C60-FF86-80ED-FBB1FAF1FE98 |
|
treatment provided by |
Felipe |
|
scientific name |
Bos gaurus Hamilton-Smith, 1827 |
| status |
|
Bos gaurus Hamilton-Smith, 1827 View in CoL
Wild Gaur
B [os]. Bubalus guavera Kerr, 1792:339 . Type locality “ Ceylon.”
Bos frontalis Lambert, 1804:57 View in CoL . Type locality “ India;” first use of the current name combination of the binomial for the domestic form ( Gentry et al. 2004; International Commission on Zoological Nomenclature 2003).
Bos gavaeus Colebrooke, 1808:512 , 516. Type locality “mountains that form the eastern boundary of the provinces of Aracan [ Burma], Chittagong (Chitgozou) [ Bangladesh], Tipara [ India], and Syilhet [ Bangladesh].”
Bos sylhetanus Cuvier, 1824:2 . Type locality “vivans à la ménagerie de Barracpour; au pied des montagnes du Sylhet.”
Bos gaurus Hamilton-Smith, 1827a:399 View in CoL . No type locality given; first use of the current name combination of binomial for the wild form ( Gentry et al. 2004; International Commission on Zoological Nomenclature 2003).
B [os. ( Bison)] gaurus: Hamilton-Smith, 1827b:373 View in CoL . Name combination.
B [os. ( Bison)] gavaeus: Hamilton-Smith, 1827b:375 View in CoL . Name combination.
Bos gour Hardwicke, 1828:231 . Type locality “Mountainous District of Ramgurh [= Ramgarh, Jharkhand, India], and Table-land of Sirgoojas [= Surguja, Chhattisgarh, India].”
Bos gayaeus Hardwicke, 1828:232 . Incorrect subsequent spelling of Bos gavaeus Colebrooke, 1808 .
Bison gaurus: Jardine, 1836a:251 View in CoL . Name combination.
Bison sylhetanus: Jardine, 1836b:257 . Name combination.
[ Bos (Bibos)] subhemachalus Hodgson, 1837:499. Type locality “saul [= sal] forest of Nipal [= Nepal].”
[ Bos (Bibos)] cavifrons Hodgson, 1837:747 Replacement name for Bibos subhemachalus Hodgson, 1837 .
Bos gareus Gray, 1843:151 . Incorrect subsequent spelling of Bos gaurus Hamilton-Smith, 1827 View in CoL .
Bos gaur Sundevall, 1844:201 . Incorrect subsequent spelling of Bos gaurus Hamilton-Smith, 1827 View in CoL .
Bibos frontatus Gray 1846:230 . Incorrect subsequent spelling of Bos frontalis Lambert, 1804 View in CoL .
[ Gaveus] frontalis: Hodgson, 1847:706 . Name combination and incorrect subsequent spelling of Bos frontalis Lambert, 1804 View in CoL .
[ Gaveus ] gayoeus: Hodgson, 1847:706. Name combination.
[ Gaveus] sylhetanus: Hodgson, 1847:706 . Name combination.
Bibos concavifrons Roulin 1849:619 . Incorrect subsequent spelling of Bibos cavifrons Hodsgon, 1837 .
Gavaeus frontalis: Horsfield, 1851:179 View in CoL . Name combination.
Bibos asseel Horsfield, 1851:181 . Type locality “South-eastern Frontier of Bengal and Silhet.”
Bos frontalis domesticus Fitzinger, 1860:387 . Name combination.
Gavaeus gaurus: Blyth, 1860:284 View in CoL . Name combination.
Bos (Bibos) frontalis: Lydekker, 1898:32 View in CoL . Name combination.
Gauribos brachyrhinus Heude, 1901:3 , 4. Type locality “ Pursat, station située sur un affluent des grand lacs du Cambodge [= on one of the tributaries of the grand lakes of Cambodia],” based on lectotype selection by Braun et al. (2001:652).
Gauribos laosiensis Heude, 1901:3 . Type locality “la chaine qui sépare le Laos du Tonkin, vers la province de Camoun [= Annamite Mountain, separating Laos and Cambodia],” based on lectotype selection by Braun et al. (2001:652).
Gauribos sylvanus Heude, 1901:4 . Type locality “foréts des Mois [= Moi forests],” Vietnam, based on holotype selection by Braun et al. (2001:652).
Gauribos mekongensis Heude, 1901:5 . Type locality “ Kratié,” Cambodia, based on lectotype selection by Braun et al. (2001:652).
Uribos platyceros Heude, 1901:5 . Type locality “Tourane de bassins des rivières de Hué,” Vietnam, based on lectotype selection by Braun et al. (2001:652).
Bubalibos annamiticus Heude, 1901:3 , 6. Type locality “Hué [Province],” Vietnam, based on lectotype selection by Braun et al. (2001:652).
Bos View in CoL ? leptoceros Heude, 1901:7. Type locality “Kampot, au bord du golfe de Siam [= Cambodia on shores of the Gulf of Siam].”
Bibos discolor Heude, 1901:3 , 8. No type locality given.
Bibos sondaicus Heude, 1901:3 , 8. No type locality given.
Bibos longicornis Heude, 1901:9 . No type locality given.
Bibos ? fuscicornis Heude, 1901:9. Type locality “Dûa sur la rivière de Vinh.”
Bos gaurus readi Lydekker, 1903:266 . Type locality “ Burma.”
Bos gaurus frontalis: Lydekker, 1912:177 Name View in CoL combination.
CONTEXT AND CONTENT. Order Artiodactyla , suborder Ruminantia , infraorder Pecora , family Bovidae , subfamily Bovinae , tribe Bovini . Lydekker (1907) classified 3 subspecies of Bos gaurus : B. gaurus gaurus ( Bangladesh, India, and Nepal), B. gaurus readei ( Burma and China), and B. gaurus hubbacki ( Malaysia) . Lydekker classified the 3 subspecies using only 5– 6 specimens, and all of the morphological differences that he relied on have since been proved to be incorrect. More recently, based on skull and horn measurements, 2 subspecies were proposed ( Groves 2003; Groves and Grubb 2011): B. gaurus gaurus in India and Nepal (and possibly Bangladesh) and B. gaurus laosiensis in Myanmar ( Burma), Lao PDR, Vietnam, Cambodia, Thailand, and West Malaysia (and presumably southern China). This new classification was also based on a small sample size of skulls. Thus far, no genetic analyses have yet conclusively corroborated the existence of subspecies. Therefore, the evidence to split B. gaurus into subspecies remains inconclusive.
NOMENCLATURAL NOTES. Although Wilson and Reeder (2005) list Bos diardii Temminck, 1838 , and Bos frontatus Temminck, 1938 , as synonyms of Bos gaurus Hamilton-Smith, 1827a , and Bos frontalis Temminck, 1838 as synonyms of Bos frantalis Lambert, 1804 —while using Bos frontalis to refer to both the wild and domestic forms—they clarify that references for both these synonyms were “nomen nudum.” Kerr (1792) included Bos bubalus guavera in his list of Mammalia of the Animal Kingdom based on the description by Knox (1681:21) of a wild buffalo with white legs called gauvera in Ceylon. Pennant (1792:31), also referring to Knox (1681), described a subspecies of buffalo from Ceylon with “legs that are white one-half way from the hoofs” as gauvera in his “History of quadrupeds.” Colebrooke (1808) provided the first detailed description and measurements of wild and domestic form and called the animal gayal, which is one of the common names for the domestic form. Hardwicke (1827) coined his Bos gour synonym based on the detailed description by Traill (1824), who described an animal he thought was known as gour in India. F. Cuvier (1824) provided the first colored illustrations of male and female B. gaurus that were based on the description of wild B. gaurus by M. Alfred Duvaucel.
Common names of the wild form in other languages include tadok (Adi); peeoug (Burmese); kaati, kaadu kona, kaadu yemme (Kannada); kulga, gameya (Kannada in Uttara Kannada district); duddu (Kannada in Northern Udupi district); meuay (Lao); seladang (Malay); Da-E-Ni, Xffi+ (Mandarin); raangawa (Marathi); kattu pothu (Malayalam); gauri gai (Nepali); kaatu maadu (Tamil); Bò Tót (Vietnamese); adavi dunna (Telugu); krating กระทง (Thai); and moo (Gonds).
DIAGNOSIS
Species in the subfamily Bovinae are large and have stout bodies, hollow horns, relatively short legs, long tails with a terminal tuft of hair, broad muzzles, and no facial, pedal, or inguinal glands ( Blanford 1888; Lydekker 1913). Five genera in Bovinae ( Grubb 2005) are currently considered in the tribe Bovini : Bison , Bos , Bubalus , Pseudoryx , and Syncerus . Species in the Bovini tribe are distinguishable by their smooth horns that are strongly keeled and spirally twisted, although not being regularly ridged ( Figs. 1–3 View Fig View Fig View Fig ; Grubb 2005; Groves and Grubb 2011). Extant species of Bovini are further distinguished from other Bovinae species by their low, wide skulls, internal sinuses in the frontals extending into the horn cores, a short braincase and widened occiput, molars with larger basal pillars and complicated central cavities, and upper molars that are strongly hypsodont ( Lydekker 1913; Gentry 1992; Grubb 2005; Groves and Grubb 2011).
Except for the yak ( Bos mutus ), which is hairy and has 14 dorsal and 5 lumbar vertebrae ( Leslie and Schaller 2009), Bos gauru s shares its traits of not being hairy and having 13 dorsal and 6 lumbar vertebrae with the other Bos and Bubalus wild cattle species of Asia. Dewlaps that hang under the neck and chin and a dorsal ridge are prominent features that distinguish Bos males from Bubalus males. Dewlaps and the dorsal ridge are prominent distinguishing features of adult male B. gaurus ( Fig. 1 View Fig ), and these traits are shared with the males of other Southeast Asian Bos species, such as the banteng ( B. javanicus ) and the now presumed extinct kouprey ( B. sauveli ). Both sexes in all 3 species have white lower legs, and females of both species are primarily brown and males are primarily black. Bos javanicus , however, has a white patch on its rump that B. gaurus does not, and reported weights suggest that B. javanicus is smaller in size than B. gaurus . Adult male B. gaurus are further distinguishable from adult male B. javanicus by their very muscular appearance, and the brown pelage of female B. javanicus is a shade lighter than the brown pelage of female B. gaurus .
GENERAL CHARACTERS
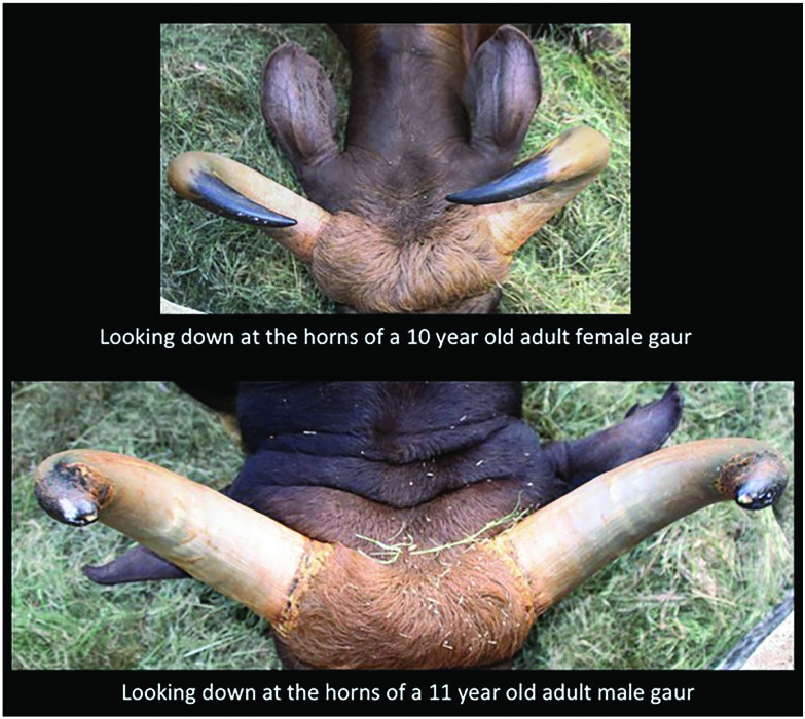
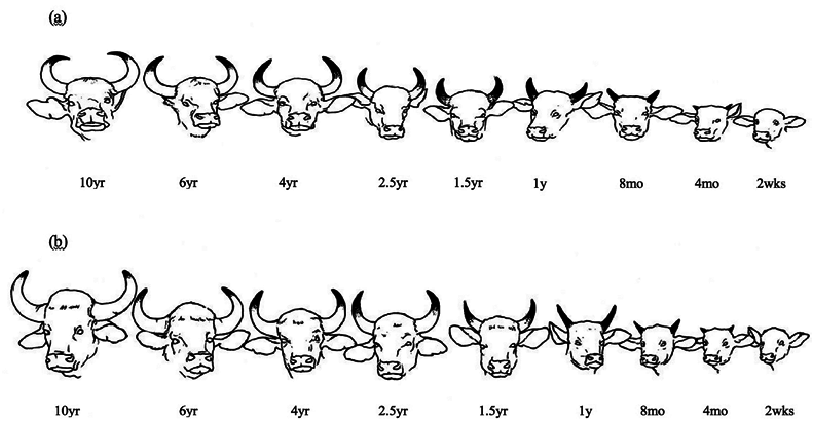
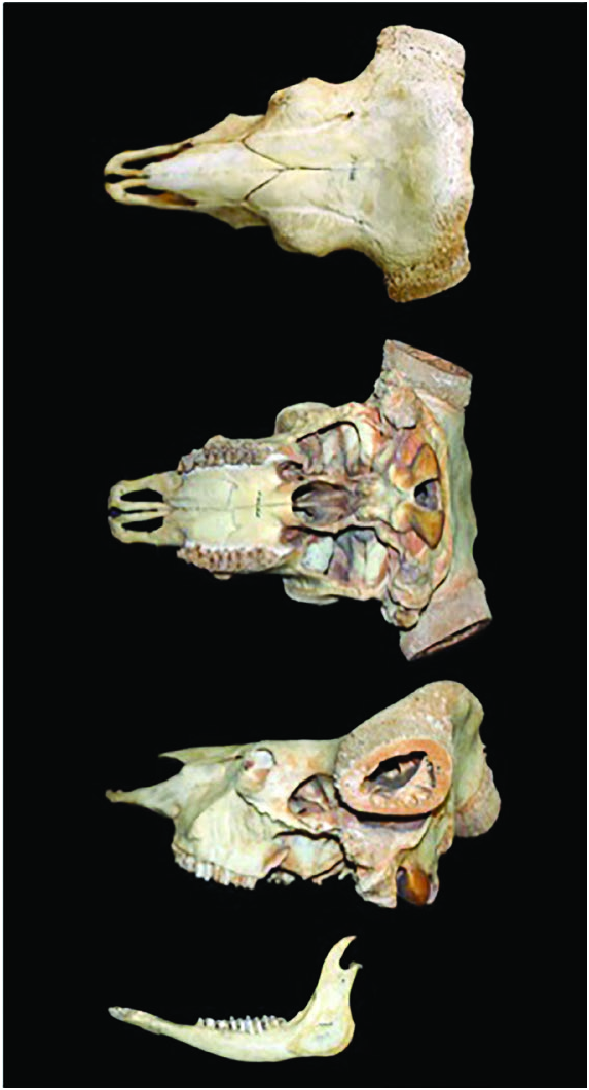
Bos gaurus is a sexually dimorphic species ( Figs. 1 View Fig and 2 View Fig ), and differences between the sexes begin to be noticeable after the age of 2 years ( Ahrestani and Prins 2011). Young calves (0–2 months) have light orange-brown body coats and do not have white stockings. The white stockings (all 4 legs, starting just above the knee in both sexes, are white) develop from the age of 3 months ( Fig. 1 View Fig ). Males grow rapidly to attain large sizes (> 900 kg) and develop a black pelage with age; adult males, generally> 5 years are referred to as black bulls. Females are smaller in size (<600 kg) and have a brown pelage ( Figs. 1 View Fig and 2 View Fig ). Both sexes have a light brown “boss” between their horns. Horns in both sexes are black in their early stages of development and, with advancing age, begin to whiten from the base up. The greater the proportion of white on the horns of a B. gaurus , the older is the individual ( Fig. 4 View Fig ). Horns of males are thicker and extend outwards first before curving inward, which results in the horns on males being further apart from each other ( Figs. 3 View Fig and 4 View Fig ). Female horns, in contrast, extend outward a lot less, and are thus closer to one another, and have a spiral curvature that makes the horns point at each other; the inward curvature of female horns begins by the age of 2 years ( Figs. 3 View Fig and 4 View Fig ). Horns on both sexes appear to grow throughout the lifetime of an individual, and it is not uncommon to see old females with horns that are nearly touching. A muscular elevated dorsal ridge and dewlaps that hang under the neck and chin easily distinguish adult males from females. Frontals and parietals of the skull are in a single plane and are similar to other Bos species ( Fig. 5 View Fig ). A more detailed description of the differences in morphological characteristics between the sexes across age classes is presented in Ahrestani and Prins (2011).
Reported weights of adult males shot in the wild were as follows: 590, 782, 864, 864, 931, and 941 kg (Dunbar-Brander 1923; Meinertzhagen 1939; Morris 1947). Reported weights of adult females shot in hunts were as follows: 440 kg (excluding blood—Schaller 1967) and 703 kg ( Meinertzhagen 1939). The skull of a male B. gaurus has been recorded to weigh about 21 kg ( Robison 1941). Based on measurements of 9 captive males and 14 captive females, Ahrestani and Prins (2011) reported a maximum shoulder height of 175 cm for males and 148 cm for females. Other records of shoulder height of males include those killed in hunts: 145 cm ( Cameron 1929), 176 cm ( Inverarity 1889), 178 cm ( Forsyth 1889), and 197 cm ( Pillay 1952). Length from nose to root of the tail of a male’s body was reported to be 249 cm ( Cameron 1929) and 284 cm (Dunbar- Brander 1923). Length of tail was 86 cm (Dunbar-Brander 1923) and 89 cm ( Cameron 1929).
The range of length of a male’s horn is 61–96 cm ( Forsyth 1889; Inverarity 1889; Baker 1903; Cameron 1929; Robison 1941; Hundley 1952; Pillay 1952; Schaller 1967), and the spread (between the widest outside points) is 89–134 cm ( Inverarity 1889; Baker 1903; Cameron 1929; Robison 1941; Hundley 1952; Pillay 1952). For males, the circumference of horns at the base has been measured to be 43–58 cm ( Baker 1903; Pillay 1952), and ear length is about 23 cm. Hind foot length is about 55 cm ( Cameron 1929); the proportion of the length of legs to its body mass is probably one of the smallest in the animal kingdom. The eyes of a B. gaurus are normally colored brown. The eyes, however, sometimes appear blue under certain light conditions because of the presence of tapetum lucidum, a membrane behind the retina that makes the eyes of several animals shine in the dark.
DISTRIBUTION
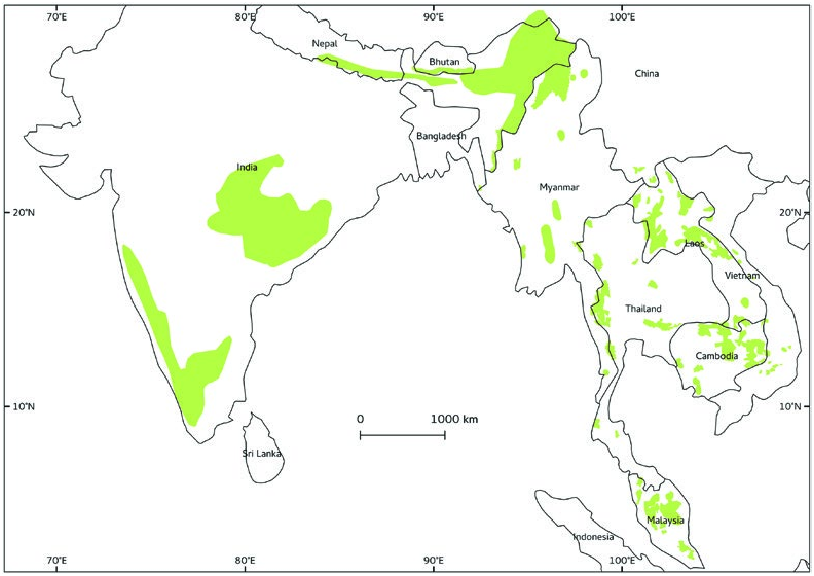
The global distribution of Bos gaurus , both historically and in present time, has been restricted to southern and southeastern Asia ( Fig. 6 View Fig ), which includes Bangladesh, Bhutan, Cambodia, China (Yunnan and southern Tibet), India, Lao PDR, Malaysia (Peninsular Malaysia), Myanmar ( Burma), Nepal, Sri Lanka (extinct), Thailand, and Vietnam. In the last century, the overall distribution of B. gaurus has shrunk by> 80%, and today, B. gaurus is mainly found in protected areas ( Schaller 1967; Choudhury 2002; Ahrestani and Karanth 2014).
Although the elevational range of habitats that B. gaurus occupies is wide, sea level to 2,700 m, it is found more on hills than on plains. Early natural history accounts of B. gaurus report that B. gaurus prefers hilly areas, particularly during the dry season. However, it is unclear if this behavior is driven by some ecological need or if it is a function of the fact that the majority of all remaining habitat available to B. gaurus is hilly terrain as nearly all suitable habitat in the plains have been lost to agriculture ( Schaller 1967).
More than 80% of the global population of B. gaurus is found in India, distributed over 3 widely separated geographical regions: the Western Ghats, central India, and northeastern India ( Choudhury 2002). The largest population of B. gaurus (3,000 –5,000) in the world is found in the 5,520 km 2 Nilgiri Biosphere Reserve, southern India. Currently, in India, populations of B. gaurus in the Western Ghats are secure; populations in central India are less secure, while populations in northeastern India are vulnerable ( Ahrestani and Karanth 2014).
Bos gaurus is most probably extinct in Bangladesh; no records have been reported since the 1970s ( Khan 1985). It is possible that individuals from Mizoram and Tripura in India occasionally cross over into Bangladesh ( Choudhury 2002). The overall population of B. gaurus in Nepal, confined mainly to Chitwan National Park and Parsa Wildlife Reserve, is understood to be less than 500 individuals, but is considered to be stable. In Bhutan, B. gaurus is apparently found all along the southern foothill zone, mostly in protected areas with a few recent sightings outside protected areas. The status of B. gaurus in Myanmar is poorly understood, though during a national tiger survey from over a decade ago, B. gaurus was camera trapped in 11 of 15 sites, with a high rate of capture in 5 of these sites ( Lynam 2003).
The overall population of B. gaurus in Thailand could be in excess of 1,000 individuals. The outlook for B. gaurus in northern Thailand is currently favorable because an effective antipoaching campaign and reforestation program over the last couple of decades has led to several increasing populations, most notably in Khao Yai National Park, Huai Kha Khaeng, and Thung Yai Naresuan wildlife sanctuaries. Forests are highly fragmented in southern Thailand, and it is assumed that B. gaurus has been largely extirpated in this region. Nevertheless, it may survive along the Malaysian border, where the human population is low and forest fragments are larger because of an ongoing insurgency in that region. Across the border, the B. gaurus population within mainland Malaysia was estimated to be around 500 in 1994; it is suspected that this population has now reduced by 50%. The outlook for B. gaurus in Malaysia is grim, and it possible that it survives at a viable population only in Taman Negara (peninsular Malaysia’s largest national park—Lynam et al. 2007).
In China, B. gaurus occurs in Yunnan and southeastern Tibet ( Ahrestani and Karanth 2014). Although the exact status of the populations in these regions is unknown, it is understood that the overall population of B. gaurus in China does not exceed 200 (H. Jianlin, pers. comm.). A report from nearly 20 years stated that B. gaurus had been extirpated from much of Yunnan province ( Xiang and Santiapillai 1993). In Cambodia, B. gaurus was widespread until the 1960s, after which the overall country population has decreased by nearly 90%. The largest population can be found in eastern Cambodia (Mondulkiri Province), and recent protection measures may have stabilized this population ( Timmins and Rattanak 2001; Tordoff et al. 2005). B. gaurus in Lao DPR was estimated to be about 1,000 individuals in the 1990s ( Byers et al. 1995); however, since then, multiple populations in Lao have been extirpated, and it is estimated that the current overall population in Lao is no more than 500 individuals. In Vietnam, the current status of B. gaurus in unknown, and it is thought that populations that remain are in serious decline. The majority of B. gaurus in Vietnam are confined to Cat Tien National Park ( Polet and Ling 2004; Nguyen 2009).
FOSSIL RECORD
The first fossil record attributed to Bos gaurus included a partial skull and horns found in the older alluvium of the Narmada (= Narbada) River in central India ( Spilsbury 1840). This fossil record, however, was not dated. In general, the origins of Bos and the relationships of fossils that have been attributed to this genus remain problematic, which when combined with the poor fossil record from Africa for the time period 7–10 million years ago has made it difficult to confirm whether or not the Bovini tribe originated in southern Asia ( Bibi et al. 2009).
Pilgrim (1939), in great detail, attributed the different Bovinae (wild cattle) fossils that had been found in India from the early 19th century to various genera, namely Proamphibos , Hemibos , Bubalus , Bucapra , Proleptobos , Leptobos , Platybos , Bison , and Bos . This splitting of Bos -like fossils into multiple genera was based mostly on partial fossils of not more than a couple of specimens found each in the Siwalik Hills ( Hemibos ); Padri, Kangra, and Hoshiarpur in the Siwaliks, Jhansi Ghat in the Narmada (= Narbada) valley, and Pemganga River in Hyderabad ( Bos ); and Pinjor in the Upper Siwaliks ( Bison and Platybos ).
In general, fossils of Bos -like species found in India have been dated as belonging to the Upper Pliocene, i.e., 3.5–5 million years ago ( Pilgrim 1939; Chauhan 2008). The majority of fossils of oxen species found in India from the late Quaternary (Pleistocene) period have been attributed mainly to Bos namadicus , the species considered to be the wild stock of the extant domestic Bos indicus ( Grigson 1985; Chauhan 2008). Fossil records of Bos namadicus that were attributed to middle–late Pleistocene (32,000 BP) were found mainly in the Godavari Valley ( Badam et al. 1984; Badam and Jain 1988). Fossils found in the Narmada valley in central India, which were also dated to the Pleistocene, were classified as belonging to Bibos gaurus ( Badam and Grigson 1990) .
Besides India, abundant skulls, horns, mandibles, and teeth of Bibos (Bos) gaurus (and supposedly also of a similar species Bibos geron ) belonging to the Pleistocene were found in different areas in China beginning in the end of the 19th century ( Colbert et al. 1953). These areas included the regions of Sichuan (= Szechuan—Matsumoto 1915), Yanjinggou (= Yenchingkou— Matthew et al. 1923; Young 1932, 1939), and the Bailong cave, Hubei ( Wang et al. 2015).
FORM AND FUNCTION
A distinguishing feature of a male Bos gaurus is its dorsal ridge, which is “formed by a row of single bones springing from the back bone immediately behind the junction of each pair of ribs, of which B. gaurus has 13 pairs” ( Inverarity 1889:295). The highest point of the dorsal ridge is above the 5th–6th rib, after which the ridge reduces in height and ends abruptly at the last rib. The dental formula is i 0/4, c 0/0, p 3/3, m 3/3, total 32 (A. Bourgeois, pers. comm.), which is the same as that of cattle. B. gaurus secretes an oily chemical through its skin, which had been noticed by early naturalists who had proposed that it acted as an insect repellent ( Hubback 1937). The oil, identified as 5-(1-hydroxynonyl)-2-tetrahydrofuranpentanoic acid, has been named bovidic acid ( Ishii et al. 2004), and 2 studies have found it to have mosquito repellent properties ( Tran and Chauhan 2007; Phillips et al. 2015).
Mean heart rates for 5 B. gaurus ranged from 49.3 to 57.7 beats/min, and mean body temperatures for 2 B. gaurus were 38.2°C and 38.8°C ( Thomas et al. 1996). Short-duration adverse stimuli caused brief 3-fold increase in heart rate, but baseline rates returned after the stressors were removed. Moving B. gaurus to novel environments or pairing them with nonaffiliates also increased heart rates ( Thomas et al. 1996). Body temperature was not affected by short-term stressors, but it was positively correlated with ambient temperature ( Thomas et al. 1996).
Bos gaurus is a ruminant and, among all extant true ruminants, is only second to the giraffe in body mass. Observations by multiple naturalists and field biologists suggest that B. gaurus depends less on sight (despite being capable of seeing at night) and more on smell and hearing to detect danger ( Inverarity 1889; Dunbar-Brander 1923; Hubback 1937; Schaller 1967; Krishnan 1972).
ONTOGENY AND REPRODUCTION
Johnston et al. (1994) found that ovarian oocytes of immature Bos gaurus were capable of in vitro maturation and fertilization with thawed homologous spermatozoa and that the resulting embryos were capable of advancing to blastocysts in culture and of producing live-born offspring after embryo transfer. Specifically, the embryos developed to the blastocyst stage in 7 days, and a live-born calf was delivered 308 days after transferring the embryos to Holstein recipients.
Primiparity (i.e., age at first birth) for captive B. gaurus is 3 years, which is similar to what has been observed for other Bovini species (Ahrestani et al. 2011). Ovulation has been found to occur at 19- to 22-day intervals, and onset of ovulation can be predicted based on measured temperature spikes ( Thomas et al. 1996). Although no data exist for B. gaurus , the average length of the estrous cycle in the domestic form Bos frontalis has been found to be 22 days, and the average duration of how long female B. frontalis remain in heat was found to be 45 days ( Giasuddin et al. 2003). Volume of semen in a single ejaculation ranges from 0.2 to 11.0 ml, and pH level in the semen ranges from 6.58 to 8.42 ( Iswadi et al. 2016).
Data from Omaha Zoo, United States, and Mysore Zoo, India, suggest that the gestation period for B. gaurus is between 9 and 10 months, about 280 days ( Ahrestani and Prins 2011). This is similar in duration to the 9-month gestation period that we know for domestic cattle and that was reported for B. gaurus by Hubback (1937). In captivity, B. gaurus has been known to give birth every year, and an 18-year-old B. gaurus was known to have given birth with no complications (Ahrestani et al. 2011). Given their 9- to 10-month gestation period, there is no reason not to assume that free-ranging B. gaurus give birth every year too (Ahrestani et al. 2011).
Schaller (1967:181) reported that “single births are the rule, there being no records of twins” for B. gaurus . Thus far, no evidence has been found to contradict this statement; no twins were recorded in 319 births of B. gaurus in European zoos until 2011 (A. Bourgeois, pers. comm.), in more than 150 births (1968–2006) in Omaha Zoo, United States, and in 41 births (1994–2006) in Mysore Zoo, India (F. S. Ahrestani, in litt.). A study of 13 B. frontalis females in Bangladesh found the mean daily milk yield and lactation length were 305 ml and 117 days, respectively ( Giasuddin et al. 2003). Although calves as old as 180–240 days have been observed suckling (F. S. Ahrestani, in litt.), it is understood that calves rarely suckle beyond the age of 6 months ( Schaller 1967). The longest continuous suckling by a calf has been recorded to be about 9 min ( Schaller 1967).
Bos gaurus appears to breed throughout the year. In a study from southern India, Ahrestani et al. (2012b) observed B. gaurus calves throughout the year, with no discernable peak, similar to what Inverarity (1889) reported for India, Peacock (1933) reported for Burma, and Weigum (1972) reported for Malaysia. There have been, however, others that have suggested mating and calving seasons in different regions. For example, Dunbar- Brander (1923) reported that the majority of mating in central India occurred in December–January, with the majority of calves born in August–September; Stebbing (1911) and Sanderson (1912) not only reported similar months but also noted that calves were born in April–June; Hubback (1937) reported seeing young in Malaysia at all times except from October to December; Morris (1937) reported that the peak rutting period in southern India was from November to March; K. U. Karanth (in litt.) found peak rutting activity in June–September (wet season) in Nagarahole, southern India; and Schaller (1967:180), while having observed calves throughout the year, reported that “the frequency of rutting call and other aspects of sexual behavior reached a peak in March and April” in Kanha, India.
ECOLOGY
Population characteristics. —The majority of density (individuals/km 2) estimates of free-ranging Bos gaurus are from India, and these estimates vary by location and time. The highest densities of B. gaurus have been found in the southern region of the Western Ghats, which include: densities as high as 10–15 in the backwaters of the Kabini dam in 2001–2002 ( Madhusudan 2004); 14.4 (± 3.8 SE) in Mudumalui Tiger Reserve in 1988–1992 ( Varman and Sukumar 1995); 12.3 (5.6–16.4, 95% CI) in the Anamalai Tiger Reserve in 2001–2004 ( Kumaraguru et al. 2011); densities in Nagarahole Tiger Reserve have been consistently high over 20 years: 5.6 (± 1.8 SE) between 1987 and 1990 ( Karanth and Sunquist 1995), 4.5 (± 0.8 SE) in the mid-1990s ( Karanth and Nichols 1998), and 5.1 (± 1.3 SE) individuals/km 2 in 2011 ( Ahrestani and Karanth 2014); 5.3 (± 1.5 SE) in Bandipur Tiger Reserve in 2011; and estimates from Bhadra Tiger Reserve in 2011 were 1.0 (± 0.4 SE) ( Ahrestani and Karanth 2014), down from 1.48 (± 0.6 SE) recorded in 2000 ( Jathanna et al. 2003). Densities of 0.7 (± 0.2 SE) in the mid-1990s ( Karanth and Nichols 1998) and densities of 0.3 (0.1–1.1, 95% CI) in 1998–1999 ( Biswas and Sankar 2002) were recorded in Pench National Park, central India. The population density in Huai Kha Khaeng Wildlife Sanctuary, western Thailand, was reported to be 1.8 (1.3–2.3, 95% CI) in 1998 ( Srikosamatara 1993).
The ratio of males:females at birth is only available for captive populations and has been recorded as 0.86 (n = 41) in Mysore Zoo, 1.36 (n = 180) in Omaha Zoo, and 1.29 (n = 16) from the National Zoological Park, Calcutta, India ( Reed 1959; Ahrestani et al. 2011). Although these reports do not provide a definite ratio, based on data of other Bovini species, Ahrestani et al. (2011) concluded that there was no reason not to assume that the sex ratio at birth for B. gaurus is parity. Given that the difference between males and females till the age of 2 years is minimal, it is not surprising that the survival rates for both sexes were found to be similar till the age of 2 years: among 109 medium-sized and large-sized calves sexed in Kanha National Park, India, 55 were male and 54 were female ( Schaller 1967); and in the Bandipur–Mudumalai landscape, India, among 282 B. gaurus sexed below the age of 1 year, 140 were males and 142 female, and among 153 B. gaurus sexed between the age of 1 and 2 years, 76 were males and 77 females (Ahrestani et al. 2011).
The ratio of adult females:males in B. gaurus populations is often 2:1, and sometimes 4:1. In southern India, the adult male:female ratio has been reported as being 25:100 ( Ashokkumar et al. 2010), 39:100 ( Ramesh et al. 2012), and 33:100 (Ahrestani et al. 2011) in Mudumalai Tiger Reserve; 18:100 ( Karanth and Sunquist 1992) in Nagarahole Tiger Reserve; 45: 100 in Parambikulam Tiger Reserve ( Vairavel 1998); in central India ratios of 60: 100 in Pench National Park ( Sankar et al. 2002) and 50:100 ( Belsare et al. 1984) and 80:200 ( Schaller 1967) from Kanha National Park; and in northeastern India a ratio of 58:100 has been reported from Trishna Wildlife Sanctuary ( Dasgupta et al. 2008). Also, survival rates of females appear to be higher than those of males. Analyzing the survival of 72 males and 58 females from birth to death in Omaha Zoo, United States, Ahrestani et al. (2011) found that after the age of 2, survival of females was higher than males in captive conditions.
Despite the difference in survival between the sexes, the maximum-recorded lifespan for both sexes in captivity has been found to be the same: about 24 years for a captive female ( Crandall 1964) and 23.6 years for a captive male (Ahrestani et al. 2011). Considering data from populations of other freeranging Bovini species, a female B. gaurus that reaches the age of 20 years may be expected to produce 8–10 calves in her lifetime (Ahrestani et al. 2011).
Space use. —Today, Bos gaurus is primarily confined to forested areas (dry and moist deciduous, semi-evergreen, and evergreen) and hilly terrain (<2,500 m) in India and across much of its distribution in Southeast Asia ( Choudhury 2002; Karanth et al. 2009; Ahrestani et al. 2012a). This appears to be a function of the forested habitat available in the protected areas to which the species is more-or-less confined; there is an acute lack of protected plains and grassland habitats in India and southeastern Asia. For example, Schaller (1967:178) suggested nearly 50 years ago that “the apparent preference of B. gaurus for hilly terrain may in part be due to the conversion of much of an earlier habitat in the plains into fields, whereas the hills have until recent years been left relatively undisturbed.” Naturalists have observed the preference for hilly terrain for more than a century ( Forsyth 1889; Inverarity 1889; Dunbar-Brander 1923), but there is no evidence to support the oft-mentioned claim that B. gaurus moves to hilly terrain to escape insects.
In Thailand, a population of B. gaurus recovering after poaching selected deciduous over evergreen forest ( Steinmetz et al. 2010). In Lao PDR, Vietnam, and Cambodia, B. gaurus is found in evergreen forests, montane forests, and in both open and closed lowland forests, with the species frequenting grassy openings in closed canopy forests ( Duckworth et al. 1999; Timmins and Rattanak 2001). In Malaysia, B. gaurus has been associated with habitats dominated by secondary vegetation, such as jungle clearings, abandoned fields of shifting cultivators, forest fringes, and openings along rivers ( Hubback 1937; Foenander 1952; Stevens 1968; Weigum 1972; Conry 1989). Multiple naturalists have reported observing B. gaurus foraging along stream banks in India ( Forsyth 1889; Inverarity 1889; Dunbar-Brander 1923; Krishnan 1972). It is possible that low-lying regions near rivers and drainage lines—areas that typically retain the last remaining herbaceous layer in the dry season—act as key foraging habitats for B. gaurus in the dry season. B. gaurus have historically and even today use mineral licks throughout their distributional range ( Inverarity 1889; Hubback 1937; Ogilvie 1954; Schaller 1967; Krishnan 1972; Weigum 1972; Steinmetz 2004).
Bos gaurus forages day and night and seems to prefer to forage at night, dawn, and dusk in areas with warm climate. During the hottest periods of the day, B. gaurus often retreats to forested areas to chew cud, often done while lying down ( Forsyth 1889; Krishnan 1972; F. S. Ahrestani, in litt.). During his seminal study, Schaller (1967) observed that the majority of B. gaurus were under shelter of trees by 0700 h, were rarely seen in the open after 0800 h, and emerged from forested patches to open grasslands in the evenings. A radio-telemetry study in Pench Tiger Reserve, India, recorded the mean daily (daytime) movement of an adult male to be 1.8 km in summer and 1.3 km in the rainy (monsoon) season and that of an adult female to be 1.2 km in the summer and 1.4 km in the rainy season ( Sankar et al. 2002).
Bos gaurus has been found to range over areas that vary from 8 to 169 km 2. A free-ranging B. gaurus herd monitored for 2 years (1967–1969) in Taman Negara National Park, Malaysia, ranged over 8 km 2, mainly along the Tembeling River ( Weigum 1972). Another telemetry study from Malaysia of 3 B. gaurus found a yearling male that ranged over 30 km 2 (November 1978 – May 1979), a yearling female that ranged over 52 km 2 (October 1977 – December 1978), and an adult male (about 5 years old) that ranged over 137 km 2 (January–November 1978; Conry 1989). Despite the availability of primary forest within 1.5 km of their home ranges, these 3 B. gaurus that were monitored in Malaysia primarily used disturbed and early seral habitats created by logging and agricultural development, secondary forest, and agricultural estates. Furthermore, areas within 500 m of agricultural fields, areas within 500 m of major rivers, and areas below 61 m elevation were used disproportionately with respect to their availability in the ranges of these individuals. These 3 B. gaurus individuals also frequently used areas adjacent to human settlements, and all their ranges included at least 1 salt lick.
In Pench Tiger Reserve, India, the ranges of an adult male and female—radio-collared and monitored for nearly a year (1997–1998)—were 12.6 and 7.3 km 2 in summer and 7.6 and 13.8 km 2 in winter, respectively ( Sankar et al. 2002). Among the 19 B. gaurus reintroduced in 2011 to Bandhavgarh Tiger Reserve, India, 3 adult males and 9 adult females were radiocollared and monitored from January 2011 to January 2012. This reintroduced herd utilized an area of 290 km 2 in summer, 137 km 2 in the rainy (monsoon) season, and 155 km 2 in winter. The ranges of individual males in this reintroduced herd varied from 135 to 142 km 2, and the ranges of individual females varied from 32 to 169 km 2 ( Sankar et al. 2013).
Bos gaurus has shown site fidelity spanning a few days to sometimes years. In Nagarahole, India, an adult female, recognized because of a horn deformity, remained within a 30 km 2 area for over a decade (K. U. Karanth, pers. comm.). It is also common to see the same solitary males in an area for months ( Inverarity 1889; F. S. Ahrestani, in litt.). B. gaurus , however, also makes local seasonal migrations: a year-long study across 200 km 2 in Bandipur, India, found B. gaurus to be confined to moist deciduous forests in the dry season and to dry deciduous forests in the wet season ( Ahrestani et al. 2012a); in both Garumara Wildlife Sanctuary, West Bengal, and the Rairakhol region, Orissa, India, B. gaurus was found to be absent during the dry season (July–October, Garumara—Guin and Pal 1982; March–July, Rairakhol—Imam 1985), but was present during the rest of the year. It is likely that such seasonal migrations are driven by B. gaurus trying to satisfy the nutritional requirements of its large body mass.
Diet. —Although Bos gaurus eats a diverse array of plant species and plant parts, a growing body of evidence suggests it is primarily a grazer. This is consistent with Hofmann and Stewart (1972) classifying Bovini species as bulk feeders of grasses, a classification that was made considering the large rumen size, the stomach structure, and the feeding habits of Bovini species such as the African buffalo Syncerus caffer and American bison Bison bison (currently recognized as Bos bison ). In general, other Bovini species in Asia that are closely related to B. gaurus , such as the water buffalo Bubalus arnee , are also considered grazers. Over a century ago, naturalists such as Inverarity (1889) claimed that B. gaurus was mainly a grazer. Since then, evidence to support this claim has grown. For example, the mean proportion of grasses found in the rumen contents of 4 B. gaurus autopsied in Kanha was 85% (range 66–100%, Schaller 1967); grasses made up 66%, browse 26%, and herbs and other plants 8% of the diet of B. gaurus studied in Nepal ( Chetri 2003, 2006); graminoids (grasses, sedges, and bamboo) accounted for> 60% of epidermal fragments found in their feces in a multisite study from central India ( Gad and Shyama 2011); and an isotopic analysis of their feces from southern India showed that B. gaurus was primarily a grazer throughout the year ( Ahrestani et al. 2012a).
Leaves and shoots of bamboo, the world’s largest grass species, are sought after by B. gaurus throughout its range: the 3 dominant bamboo species in southern India ( Bambusa arundinacea , Dendrocalamus strictus , and Oxytenanthera monostigma ) are all eaten ( Krishnan 1972; K. U. Karanth, pers. comm.), and in Thailand, B. gaurus has been found to preferentially eat bamboo shoots in the wet season and bamboo leaves in the dry season ( Prayurasiddhi 1997; Steinmetz 2004).
Bos gaurus , however, eats far more species than just grasses. For example, Gad and Shyama (2011) found that B. gaurus fed on 7 grass species, 5 herb species, 8 shrub species, and 12 tree species; in Nepal, B. gaurus has been found to feed on 49 plant species (23 grasses, 17 browse species, and 9 herbs and others— Chetri 2003, 2006) and in Malaysia 89 different plant species, of which only 38 were grass species were reported ( Weigum 1972); in Kanha, B. gaurus have been found to feed on 7 shrub species, 6 grass species, 3 vine species, 4 forb species, fruits from 2 tree species, and leaves from at least 17 tree species ( Schaller 1967); and Dunbar-Brander (1923) recorded B. gaurus feeding on more than 40 plant species. In some areas, the proportion of browse in their diet might vary by season; for example, in a 4-year study, Gad and Shyama (2011) found that the proportion of browse consumed by B. gaurus increased from 15% in winter to 40% in summer.
Although B. gaurus feeds on different plant parts, it primarily eats leaves. For example, in their histologic study, Gad and Shyama (2011) found that 87% of B. gaurus diet composed of leaves. B. gaurus feeds primarily by extending its tongue to curl around leaves and twigs, which it pulls into its mouth and bites off with a tug ( Schaller 1967; Krishnan 1972). Multiple studies have found B. gaurus feeding on fruits (Dunbar-Brander 1923; Schaller 1967; Krishnan 1972; Chetri 2003), and its eating of shoots is generally associated with its foraging on bamboo ( Forsyth 1889; Krishnan 1972; Gad and Shyama 2011). B. gaurus has also been observed eating the bark of trees, including Adina cordifolia (Dunbar-Brander 1923) , teak Tectona grandis ( Pasha et al. 2002) , and cashew Anacardium occidentale ( Gad and Shyama 2011) in India and Holarrhena antidysentrica and Wendlandia natoniana in Malaysia ( Ogilvie 1954). The eating of bark, which has often been recorded in summer, is understood to be in response to either a shortage of forage or mineral resources, or both.
Diseases and parasites. —The cattle diseases rinderpest, foot-and-mouth, and anthrax have been reported as affecting Bos gaurus populations from the very earliest reports of the species. Rinderpest has been the most common and widespread of the 3 diseases: rinderpest was reported to have affected B. gaurus populations in India as early as the 19th century ( Inverarity 1889; Baker 1890); the disease affected populations in central India in the early part of the 20th century (Dunbar- Brander 1923; Stewart 1927); and Schaller (1967:181) reported that “a virulent epidemic of rinderpest killed many B. gaurus in the Kanha Park area in the years 1925–26.” In southern India, rinderpest has been reported to have killed hundreds of B. gaurus in the 1st one-half of the 20th century ( Anderson 1954), to have all but wiped out B. gaurus from Mudumalai in 1968 ( Krishnan 1972), and to have nearly wiped out the B. gaurus population in Bandipur in 1989 (D. V. Girish, pers. comm.). In northeastern India, rinderpest epidemics killed several hundred B. gaurus in Raimora, Assam, in 1967, in the Rairakhol region, Orissa, in 1972, and in Berbera and Dhuanali, Orissa, in 1973 ( Imam 1985). Although epidemics of foot-and-mouth disease have never been reported for B. gaurus populations, Morris (1949) reported shooting an adult male that had foot-and-mouth disease; a case of foot-and-mouth disease was reported from Hyderabad, southern India ( Ali 1953); 4 Bos frontalis contracted foot-and-mouth disease and died within 10 days in October 1990 in Calcutta Zoo ( Choudhury 2002); in March 2007, a male afflicted with foot-and-mouth disease was observed in Bandipur (F. S. Ahrestani, in litt.); and 2 B. gaurus died of foot-and-mouth disease in Bannerghatta Park, India, after a major outbreak of the disease in domestic cattle in neighboring villages ( Chandranaik et al. 2015). Thus far, there has been only 1 report of a B. gaurus dying from anthrax ( Peacock 1933).
Mycobacterium paratuberculosis was found in “histopathological sections of the intestine mucous” during the autopsy of an adult male B. gaurus that fell ill and subsequently died in Palamau Tiger Reserve ( Lal and Ashraf 1993). In Kanha, a B. gaurus female autopsied was lightly infested by nematodes ( Oesophagostomum radiatum ) in the large intestine and trematodes ( Gastrothylax crumenifer ) in the rumen, and ticks ( Boophilus microplus ) were found on 2 adults that were checked for ectoparasites ( Schaller 1967). Histologic examination of 2 B. gaurus that died in the Oklahoma City Zoo, United States, showed that they had Sarcosporidiosis, an infection caused by the intracellular protozoan parasite Sarcocystis ( Welch and Zimmer 1981) .
Interspecific interactions. —There have been no reports of Bos gaurus fighting with other herbivores, and in general, B. gaurus is considered to be tolerant of other herbivores. While old bachelor males are tolerant of the presence of humans, females and herds are generally shy of humans, avoiding contact as much as possible. There have been, however, reports of humans being charged by B. gaurus , and even fatalities from such attacks. Reports of such encounters have been related to B. gaurus charging after being shot or wounded ( Forsyth 1889), after being inadvertently surprised by people in forests ( Inverarity 1889; Morris 1952, 1953), after being injured by tigers (K. U. Karanth, pers. comm.), and when disturbed by close-up flash photography (F. S. Ahrestani, in litt.).
Bos gaurus calves are preyed on by leopards Panthera pardus , dhole Cuon alpinus , and tigers, although adults are preyed on primarily by tigers (Ahrestani et al. 2011). Leopards and tigers typically kill calves and females by asphyxiation using bites to their throats; 88% of 33 B. gaurus that were recorded killed by tigers were found with bites to their throats in Nagarahole, India ( Karanth and Sunquist 1995). Males, however, have a thick neck and often a dewlap, which makes killing them by asphyxiation difficult. Tigers have been known to overcome this by biting the hock, thereby hamstringing and bringing down male B. gaurus (Dunbar-Brander 1923) .
Records of tigers preying on calves and adults, including solitary males, date back to nearly a century ago (Dunbar-Brander 1923). Reports from different areas suggest that this behavior has not changed: the cause of death of B. gaurus in Kanha in 1964 was attributed mainly to tiger predation ( Schaller 1967); a carcass of an adult female B. gaurus killed by a tiger, and the remains of B. gaurus were regularly found in tiger feces within Eravikulam National Park, India ( Rice 1986); analysis of scats of the primary predators in Mudumalai Tiger Reserve, India, showed that B. gaurus made up 0.19% of dhole diet and about 5% of tiger diet ( Johnsingh 1983); and adult female and adult male B. gaurus comprised 23% and 15% of tiger kills, respectively, in Nagarahole, India ( Karanth and Sunquist 1995, 2000).
Bos gaurus has been observed facing the direction of a perceived threat with a raised head, raised muzzle, and flared nostrils ( Schaller 1967; Krishnan 1972; Belsare et al. 1984; F. S. Ahrestani, in litt.). In such situations, males and females often emit short, deep, and loud grunts and snorts that have been described as follows: “a violent and rapid expulsions of air through the nostrils” ( Hubback 1937:275); “a trumpetlike blast of air through the nose, accompanied occasionally by a growling sound—a harsh, rolling bru-u-u-u” ( Schaller 1967:186); and “a call spelled pff-hong; the pff is the noise made by the rush of air past the lips before the note is struck” (Dunbar-Brander 1923:151). These snorts or grunts often “raise every head in the herd” (Dunbar-Brander 1923:151; F. S. Ahrestani, in litt.). If a herd has calves, the females in the herd are known to bunch together in a muskox fashion, i.e., backs to each other and facing outwards, and have been observed leaping and kicking their hind legs, snorting and tossing their horns, and holding their heads low and advancing, sometimes rapidly toward detected leopards and tigers and other perceived threats ( Krishnan 1972; Johnsingh 1983; Karanth 1984; Tyabji 1989; F. S. Ahrestani, in litt.).
When fleeing from danger, which it often does, B. gaurus has been observed giving “a series of rather stiff-legged bounds, some 2 to 8 in number, with their forelegs brought down hard in unison to produce a series of distinct thumps. After thumping the ground, they usually trot off without making further sounds other than the usual commotion of a large animal moving through the forest” ( Schaller 1967:186). Finally, males are capable of killing tigers: corpses of a dead tigress and a male were both found after what was evidently a fight to death ( Blackburn 1934), and a radio-collared tiger that was found dead was apparently killed by a B. gaurus (K. U. Karanth, pers. comm.).
HUSBANDRY
The mithun/gayal/mithan Bos frontalis , the domestic form of Bos gaurus , is found in northeastern India, Bhutan, Myanmar, and China. B. frontalis looks very similar to B. gaurus : the males have small dewlaps and a pronounced dorsal ridge, and both sexes have white lower legs. However, B. frontalis differs from B. gaurus by being slightly smaller in size and by the shape of its horns, i.e., horns of both male and female B. frontalis extend outwards and are straight, with only a gentle curve inwards. In contrast, the horns of female B. gaurus barely extent outwards and curve significantly inwards, and though the horns of male B. gaurus extend outwards, they curve noticeably inwards at their ends. The domestication of B. frontalis might be as old as the Indus Valley civilization (Clutton-Brock 1987), but B. frontalis are rarely milked, are rarely used as draught or plough animals, and are often allowed to range free, though some choose to return to village pens for the night. B. frontalis is primarily used as a status symbol, to barter for goods, pay for brides, and is often eaten after being sacrificed for various occasions, including weddings, burials, prayers to ward off misfortune etc.
Bos gaurus males have been found mating with B. frontalis females, and there have been reports of B. frontalis × B. gaurus hybrids ( Baker 1890; Gee 1964; Simmons 1984). Beginning in 1983, a B. gaurus male was interbred with Sahiwal Friesian dairy heifers on a Malaysian Government, Department of Veterinary Services farm, and these hybrids outgrew other dairy calves ( Kamalludin 2009). B. frontalis and cattle have been deliberately interbred in Bhutan, and all cases of interbreeding between B. gaurus and B. frontalis and between B. gaurus and cattle have resulted in only a few fertile females and always in infertile males ( Simmons 1984).
Initial attempts to breed B. gaurus in captivity failed ( Forsyth 1889). The first successful and consistent breeding of B. gaurus occurred at the National Zoological Park, Washington, D.C., United States, when a male and female—obtained from Mysore, India, in 1937—produced 13 offspring (1940–1957) till the male died ( Crandall 1964). Since then, there have been over 319 B. gaurus births in European zoos, an equal number have been born in US zoos, and there have been over 50 births in Indian zoos. Today, the largest captive B. gaurus population is in Omaha Zoo (there over 70 individuals housed in 2 subpopulations), and across India, there are over 70 in captivity. Many captive populations have a high degree of inbreeding. For example, all 23 B. gaurus that were present in Mysore Zoo in 2005 were bred from 3 founder individuals that were captured 15 years ago in the wild. The male:female ratio in inbred captive populations has been skewed toward males, 62:38, in contrast to the 46: 54 male:female ratio found in captive populations that were not inbred ( Hintz and Foose 1982).
The 1st group of Bos gaurus individuals relocated from Kanha Tiger Reserve to Bandhavgarh Tiger Reserve, India, in 2011 were immobilized before relocation using a combination of etorphine hydrochloride and azaperone, and the 2nd group of individuals relocated in 2013 were immobilized before relocation using a combination of thiafentanil and ketamine ( Nigam et al. 2014; P. Nigam, in litt.).
BEHAVIOR
Grouping behavior. — Bos gaurus is predominantly a herding animal ( Fig. 7 View Fig ). Females of all ages and males below the age of 3 years are nearly always found in herds, and adult males are found both within herds and alone ( Inverarity 1889; Hubback 1937; Morris 1937; Schaller 1967). Herds of only adult males are uncommon, though in 2007, 16 males (> 4 years) were once observed together in Mudumalai, India (F. S. Ahrestani, in litt.). Given the absence of detailed studies of B. gaurus , it is unclear how herds are formed, maintained, change over time, are organized socially and hierarchically, and how individuals in a herd are related to each other.
The majority of solitary males are old, black bulls, which suggests that the solitary nature of males increases with advancing age. As solitary adult males appear to be fearless of man, and mostly anything else too, they are a common sight in areas with high densities of B. gaurus . For example, 137 of 385 (35%) encounters of B. gaurus over a 12-month period (2006–2007) in the Bandipur–Mudumalai landscape were of solitary adult males (F. S. Ahrestani, in litt.), and in other studies from Mudumalai, 48% of males observed in 2008–2009 ( Ramesh et al. 2012) and 52% of all B. gaurus observed in 1976–1978 were solitary adult males ( Johnsingh 1983).
Black bulls, however, are not always solitary. Of 200 herds tallied in Kanha, about 50% had a black bull ( Schaller 1967), and 56 of 147 herds encountered in the Bandipur–Mudumalai landscape had a black bull (F. S. Ahrestani, in litt.). Herds often include 2–3 black bulls and sometimes even include 6– 8 adult males ( Schaller 1967; F. S. Ahrestani, in litt.). The structure of herds with adult males, especially black bulls, is most probably fluid; there is, however, not enough information to understand when adult males join herds, how long they remain in a herd, when they leave a herd, and how such behavior changes with age.
Mean herd size is about 6–7 and varies by study and location: 7.8 (± 0.51 SE) in Mudumalai, India ( Ashokkumar et al. 2010; Ramesh et al. 2012); 4 (± 0.21 SE) in Trishna Wildlife Sanctuary, India ( Dasgupta et al. 2008); 4.6 (± 0.29 SE) in Pench Tiger Reserve, India ( Sankar et al. 2002); 6 in Parambikulam Tiger Reserve, India ( Vairavel 1998); 6.9 in Nargarahole Tiger Reserve, India ( Karanth and Sunquist 1992); 6 in Palamau Wildlife Sanctuary, India ( Sahai 1977); 6.5 ( Belsare et al. 1984) and about 9 ( Schaller 1967) in Kanha Tiger Reserve, India; 5.5 (± 5.2 SD) in Vietnam ( Nguyen 2009); about 11 in Malaysia ( Hubback 1937); and 10–20 in Burma ( Peacock 1933). Large (> 50 individuals) aggregations of B. gaurus have also been observed ( Sanderson 1912; Mustill 1938; Johnsingh 1983; F. S. Ahrestani, in litt.). These large aggregations appear to be temporary composite herds, i.e., multiple smaller herds coming together to form 1 large herd, most likely to exploit a favorable forage resource in an area.
Although understood to be an uncommon occurrence, males do fight each other ( Inverarity 1889; Krishnan 1972; Schaller 1967). One such confrontation involved a pair of males fighting next to a B. gaurus herd, with the pair displaying at each other, thumping the ground and snorting repeatedly, the younger of the 2 repeatedly threatening the older combatant with horns held low, and the 2 males making contact with their heads multiple times, pushing against each other, and once twisting their entwined horns ( Johnson 1986). The confrontation finally ended by the older male managing to push the younger male down a slope, and the younger male was found limping a couple of days later. Another report describes “2 bison fighting, shoving each other about, and when disengaged, swinging their heads from side to side with a twisting motion, and in so doing giving each other terrific blows on the horns” (Dunbar-Brander 1923:150). Schaller (1967) observed that the bony boss (the forehead between the horns) of the males absorbed much of the headbutting in male–male confrontations.
Adult males have been observed making a lateral (oblique) end-on display, which appears to be a nonphysical method to establish dominance ( Krishnan 1972). This display, first described by Schaller (1967:192), is “when a bull B. gaurus is seen standing still with its feet together and back held somewhat hunched, in the presence of another bull.” Such lateral displays can last several minutes. Adult males also threaten each other by lowering their heads and walking up to opponents, laterally sweeping their horns, emitting low moans and grumbles, snorting, thumping the ground with hooves, and sometimes running or walking around in circles while swiping at undergrowth with their lowered horns (Dunbar-Brander 1923; Krishnan 1972; Belsare et al. 1984; F. S. Ahrestani, in litt.). Although older black bulls do not commonly spar with each other, younger (3–5 years) adult males spar by facing each other with lowered heads ( Fig. 8 View Fig ), locking horns, and then twisting their locked heads from side to side ( Schaller 1967; F. S. Ahrestani, in litt.). Schaller observed 81 sparring incidents during his study, the majority of which (48%) were between young adult males, 10% were between young adult males and adult females, and 14% were between adult females. There are also reports from over a century ago of adult males sparring with their horns and butting heads ( Inverarity 1889).
Reproductive behavior. —Adult males and females have been observed licking each other, and it is assumed such behavior is related to courtship. Schaller (1967:198) reported that “cows and bulls lick each other’s necks, shoulders, and rump, occasionally for as long as 10 min without interruption, and bulls also lick each other at times, in 5 out of 7 instances the subordinate individual licked the individual of higher rank.” B. gaurus males exhibit flehmen, i.e., raising the muzzle and curling the lips after licking a female’s vulva, or sniffing a female’s urine or feces, a habit that is common among ungulates ( Schaller 1967). Males have also been observed tending to females—a behavior originally described for American bison ( McHugh 1958) —which is when a male stands beside a female while she grazes and then follows her step for step as she moves and grazes ( Schaller 1967).
Females separate from their herd to give birth alone and have been known to stay away for as long as 4 days ( Sanderson 1912). Mothers tend to thoroughly lick calves directly after their birth ( Krishnan 1972). Calves below the age of 2 months have been found alone lying motionless in undergrowth with their necks stretched out close to the ground that appear to be attempts to remain concealed ( Inverarity 1889; Hubback 1937; Ahrestani et al. 2011). Calves and yearlings (<18 months) are always seen in herds with adult females, and a young calf (<3 months) is nearly always found at the heels of an adult female, presumably its mother.
Communication. —Older (black) bulls make a call, presumably a rutting call, that is so unique that it distinguishes an adult male Bos gaurus from other animals as much as its other prominent features do, such as the dorsal ridge and dewlap. When this call is heard, it is common to hear other black bulls responding with the same call. The black bull begins the call by keeping its head low, and as the call progresses, it raises its head slowly and stretches out its neck, pointing its muzzle upwards with lips partly open and the whites of its eyes showing with the effort. The call has been described by many naturalists in their own words: “as the most absurd piping or whistling sound, more like the call of a bird than anything else, and absurd because such a strange sound emanates from an animal so large and powerful” (Dunbar-Brander 1923:151); “a most peculiar sound, a cross between the bugling of a wapiti (elk) and the trumpeting of an elephant, but at the same time a melodious sound that carries a long way” ( Hubback 1937:275); “a song, because of its musical qualities that is not low but can be heard a mile away” ( Schaller 1967:195); and “a long drawn, resonant, low and high pitched at the same time, not necessarily loud if heard at close quarters, but has the ability to be heard one-half a mile away” ( Krishnan 1972:335). The call has been described in detail as follows: “a clear, resonant u-u-u-u-u about one to 3 seconds long, either constant in pitch or slightly rising and falling; this note may be followed by a second one somewhat lower in tone, by a third one still lower, and so forth, giving the impression of someone practicing musical scales; and as many as ten seconds sometimes elapse between the 1st and last note ( Schaller 1967:195).” In contrast to adult males, females, besides snorting during fear and threatening behavior, are only known to moo, sounding just like cattle (Dunbar-Brander 1923; Krishnan 1972; F. S. Ahrestani, in litt.). It is still unclear, however, if the mooing is a mother communicating to its calf or is communication for some other reason.
Miscellaneous behavior. —Similar to cattle, Bos gaurusgrooms itself by licking its hide; rubbing its neck, rump, and sides against trees; or scratching itself with the tips of its horns. When B. gaurus lies down, it generally does so with its legs folded under its body, but it will occasionally also lie flat on its side with legs stretched out. B. gaurus generally lies down under the shade of trees and within swards of tall grass, where it is difficult for it to be detected in the flickering shadow of trees ( Forsyth 1889).
GENETICS
The amplification of the cytochrome b (Cyt b) gene, used as a mitochondrial DNA genetic maker, produced a 154- and 603-bp fragment in the DNA sequence of a Bos gaurus from Malaysia ( Romaino et al. 2014). Testing 130 cattle microsatellite markers on a panel of 11 individual B. gaurus from Vietnam found amplification of 117 markers (90%) with a total of 264 alleles ( Nguyen et al. 2007). Of the 117 makers, 68 were polymorphic that had 2–6 alleles per locus, and 3 cattle Y chromosome microsatellite markers (INRA124, INRA126, and BM861) were specific to B. gaurus ( Nguyen et al. 2007) .
Sequencing the entire genome of an individual, the domestic form Bos frontalis detected 23,828,562 single-nucleotide polymorphisms (SNPs) and identified 16,901 breed-specific nonsynonymous SNPs among 6,167 genes ( Mei et al. 2016). Annotation of these SNPs showed that 78.2% of the SNPs were located in intergenic regions; 21.1% were located in genic regions, including intronic regions, splicing sites, exonic regions, and untranslated regions; and the remaining 0.7% were located in upstream or downstream regions. This sequencing found the homozygous/heterozygous ratio to be 1:0.8, and Nei’s unbiased mean heterozygosity and the mean allele number across loci were 0.23 and 2.2, respectively ( Mei et al. 2016). Another cytogenetic study of the domestic form B. frontalis found the karyotype of the female B. frontalis (n = 4) comprised 58 chromosomes, including 54 acrocentric and 4 large submetacentric chromosomes ( Qu et al. 2012).
A genetic analysis of a B. gaurus female at Toronto Zoo detected a chromosome anomaly in an individual that had 2 n = 57 chromosomes (an extra submetacentric, but 2 less acrocentric chromosomes) instead of the normal 2 n = 58 found in B. gaurus , which consists of 27 pairs of acrocentric chromosomes, 1 pair of submetacentric chromosomes, and the submetacentric sex chromosomes ( Mastromonaco et al. 2004).
The genotype data collected from 117 successfully amplified microsatellites used to assess the genetic diversity within the reaming Bos gaurus population in Vietnam—estimated to be no more than 500 individuals—found the mean polymorphic information content (PIC) to be 0.252 (range: 0.083 – 0.767) and mean heterozygosity (Ho) to be 0.269 (range: 0.091 –0.909 — Nguyen et al. 2007). Multiple studies in the last decade have genetically investigated whether B. frontalis is a domestic version of B. gaurus , and the majority of these studies have found evidence that this is indeed the case. These studies include sequencing the Cyt b genes of 33 B. frontalis from Myanmar and Bhutan and finding B. gaurus haplotypes in 28 of these individuals ( Tanaka et al. 2011); sequencing the 16S rRNA gene in the mitochondrial DNA of mithun from Bhutan that demonstrated phylogenetic proximity to B. gaurus ( Dorji et al. 2010) ; and sequencing the Cyt b genes of 11 gayal from Yunan, China, found 6 haplotypes that clustered around B. gaurus and other domestic cattle species such as Bos taurus and Bos indicus ( Li et al. 2008) . In contradiction to the above studies, a study that sequenced the Cyt b genes from 28 B. frontalis from Yunnan, China, 13 B. frontalis from Arunachal Pradesh, India, and 1 B. gaurus from Yunnan, China, found the genome sequence of B. frontalis to differ from the B. gaurus ( Baig et al. 2013) . This individual study, however, does not provide enough conclusive evidence to override all the other genetic evidence and the detailed morphological comparisons that collectively strongly suggest that the B. frontalis is indeed the domestic derivative of B. gaurus .
Bos gaurus is one of the first mammals to have been cloned. Somatic cells from the skin of a male were successfully electrofused with enucleated oocytes from domestic females ( Lanza et al. 2000). One of these embryos successfully developed in a surrogate domestic female. The calf that was successfully delivered, however, developed a fatal bacterial infection 2 days after birth and died on 8 January 2001.
CONSERVATION
The global Bos gaurus population is estimated to be 13,000 – 30,000 and is projected to decline by 30% over the next 3 decades ( Duckworth et al. 2016). B. gaurus is listed as “Vulnerable” by the International Union for Conservation of Nature, and its greatest threats are loss of habitat, being poached for its meat and horns, and contracting fatal diseases from overlapping cattle populations. Country-wide populations in Bangladesh, Cambodia, China, Laos, Malaysia, Vietnam, and Thailand have declined by over 70% in the last 2–3 decades, although the decline in India and Nepal have been considerably lower. Recent poaching, mainly for meat and horns, has decimated the B. gaurus population in Malaysia, and there is a fear that the hunters responsible for this catastrophe could be soon targeting populations in other countries. Conservation action in Thailand has recently shown that it is possible to stall the decline of B. gaurus populations prevalent across southeastern Asia. On a positive note, multiple protected areas in southern India have recorded increasing trends in their B. gaurus populations over the last couple of decades.
In 1995, the last remaining herd of B. gaurus left Bandhavgarh Tiger Reserve. To restore B. gaurus in Bandhavgarh, 19 individuals in January 2011 — 5 males (3 subadults, 2 adults) and 14 females (1 yearling, 5 subadults, and 7 adults)—were translocated from Kanha Tiger Reserve to Bandhavgarh. Since the reintroduction, the newly established B. gaurus population in Bandhavgarh has been thriving and multiplying.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bos gaurus Hamilton-Smith, 1827
| Ahrestani, Farshid S 2018 |
Bos gaurus frontalis
| : Lydekker 1912 |
Bos gaurus readi
| Lydekker 1903: 266 |
Gauribos brachyrhinus
| Heude 1901: 3 |
Gauribos laosiensis
| Heude 1901: 3 |
Gauribos sylvanus
| Heude 1901: 4 |
Gauribos mekongensis
| Heude 1901: 5 |
Uribos platyceros
| Heude 1901: 5 |
Bubalibos annamiticus
| Heude 1901: 3 |
Bibos discolor
| Heude 1901: 3 |
Bibos sondaicus
| Heude 1901: 3 |
Bibos longicornis
| Heude 1901: 9 |
Bos (Bibos) frontalis:
| Lydekker 1898: 32 |
Bos frontalis domesticus
| Fitzinger 1860: 387 |
gaurus
| : Blyth 1860: 284 |
frontalis:
| Horsfield 1851: 179 |
Bibos asseel
| Horsfield 1851: 181 |
Bibos concavifrons
| Roulin 1849: 619 |
Gaveus] frontalis:
| Hodgson 1847: 706 |
Gaveus
| Hodgson 1847 |
Gaveus] sylhetanus:
| Hodgson 1847: 706 |
Bibos frontatus
| Gray 1846: 230 |
Bos gaur
| Sundevall 1844: 201 |
Bos gareus
| Gray 1843: 151 |
Bos (Bibos)]
| Hodgson 1837: 499 |
Bos (Bibos)] cavifrons
| Hodgson 1837 |
Bibos subhemachalus
| Hodgson 1837 |
Bibos cavifrons
| Hodsgon 1837 |
Bibos
| Hodgson 1837 |
Bison gaurus:
| Jardine 1836: 251 |
Bison sylhetanus:
| Jardine 1836: 257 |
Bos gour
| Hardwicke 1828: 231 |
Bos gayaeus
| Hardwicke 1828: 232 |
Bos gaurus Hamilton-Smith, 1827a:399
| : Hamilton-Smith 1827: 399 |
Bison)] gaurus
| : Hamilton-Smith 1827: 373 |
Bos gaurus
| : Hamilton-Smith 1827 |
Bison)] gavaeus:
| Hamilton-Smith 1827: 375 |
Bos gaurus
| : Hamilton-Smith 1827 |
Bos gaurus Hamilton-Smith, 1827
| : Hamilton-Smith 1827 |
Bos gaurus Hamilton-Smith, 1827
| : Hamilton-Smith 1827 |
Bos sylhetanus
| Cuvier 1824: 2 |
Bos gavaeus
| Colebrooke 1808: 512 |
Bos gavaeus
| Colebrooke 1808 |
Bos frontalis
| Lambert 1804: 57 |
Bos frontalis
| Lambert 1804 |
Bos frontalis
| Lambert 1804 |
Bubalus guavera
| Kerr 1792: 339 |
Bos
| Linnaeus 1758 |