
Veromessor chicoensis, Smith, 1956
|
publication ID |
https://doi.org/10.11646/zootaxa.5206.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:CE749F6C-5832-4152-AB4B-6D89ACCDD560 |
|
DOI |
https://doi.org/10.5281/zenodo.7327842 |
|
persistent identifier |
https://treatment.plazi.org/id/039387FD-FFC5-FFE8-7FC5-FF1B58B4FC65 |
|
treatment provided by |
Plazi |
|
scientific name |
Veromessor chicoensis |
| status |
|
( Figures 1C View FIGURE 1 , 6A View FIGURE 6 , 15 – 18 View FIGURE 15 View FIGURE 16 View FIGURE 17 View FIGURE 18 )
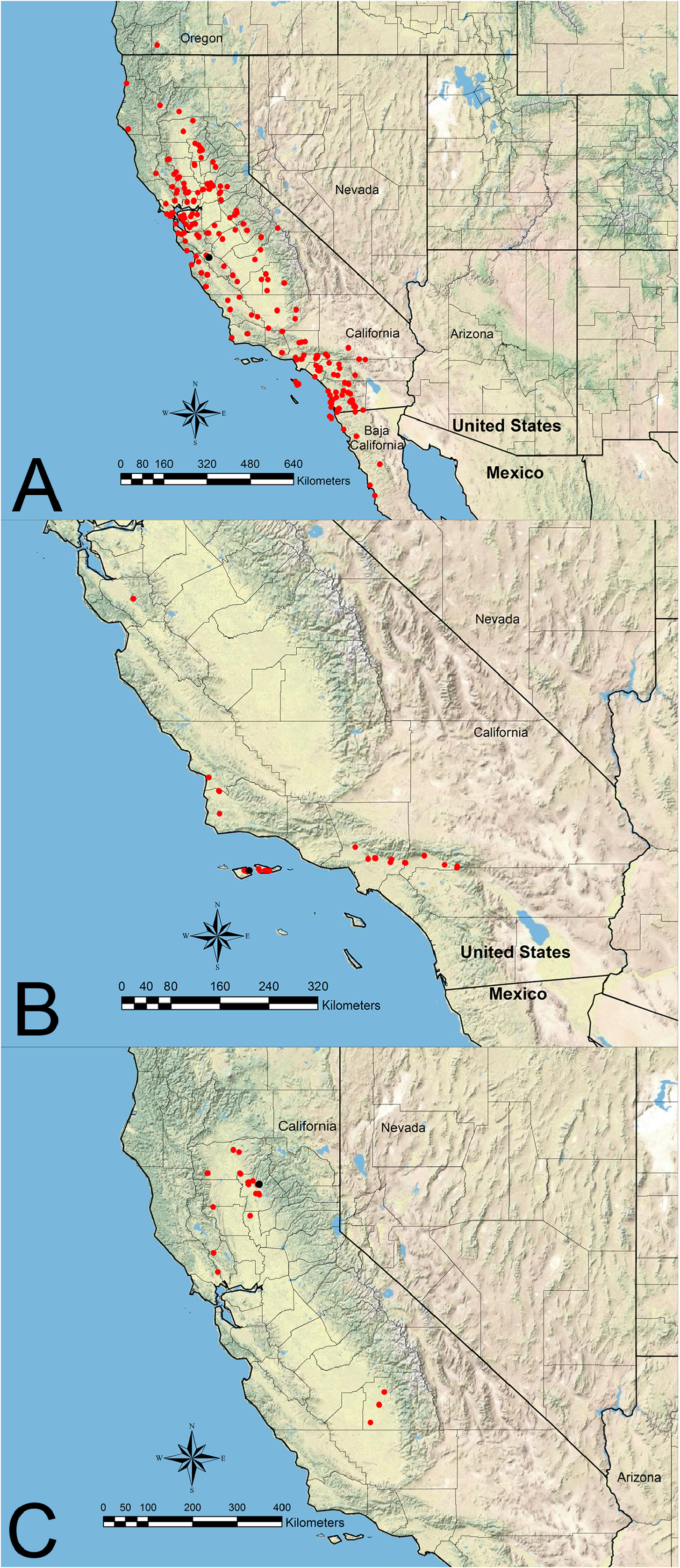
Distribution— Figure 10C View FIGURE 10
Veromessor stoddardi subsp. chicoensis Smith, 1956: 37 (worker). Types examined: holotype worker [USNM: USNMENT00529087], USNM#62959, 8 paratype workers [ USNM], 6 paratype workers [ LACM], UNITED STATES , California: Butte County, Chico (T22N, R2E, Section 17), 450’, 2 March 1954 (Adrian Wenner).
Messor stoddardi subsp. chicoensis (Smith) View in CoL ; Bolton, 1995: 253 (first combination in Messor View in CoL ).
Messor chicoensis Smith ; Johnson, 2000: 96 (raised to species, in list); see also Ward, 2005: 13 (in list).
Veromessor chicoensis Smith ; Ward, Brady, Fisher, and Schultz, 2015: 13 (revived combination in Veromessor ).
Worker diagnosis. This species is uniquely characterized by the following combination of features: (1) head and mesosoma orangish-red or orangish-brown to reddish-brown, gaster blackish-orange to blackish, (2) medial lobe of clypeus lacking medial groove but with several lateral longitudinal rugae, medial lobe not thick and protuberant in profile, not elevated above lateral lobes in frontal view, (3) mandibles with 8 teeth, (4)scape narrowest near midlength, (5) MOD distinctly less than OMD, OI <23.0, (6) cephalic dorsum weakly shining between fine longitudinal rugae, usually becoming somewhat reticulate on posterior margin; antennal fossa sometimes with several concentric rugae, (7) psammophore poorly developed; ventral surface of head capsule with scattered straight or evenly curved hairs, but J-shaped hairs mostly absent, (8) pronotum with fine, wavy to irregular, longitudinal rugae; sides of pronotum with longitudinal rugae that sometimes fade to disappear posterad, becoming granulate posterad; mesonotum with wavy to weakly irregular longitudinal rugae; mesopleura with wavy to irregular rugae that traverse longitudinally to posterodorsally, rugae sometimes with lateral branches, interrugae moderately to strongly granulate, (9) propodeal spines short, acuminate, length less than distance between their bases; infraspinal facet and propodeal declivity strongly coriarious or rugose, weakly dull to weakly shining, and (10) metasternal process large, higher than long with broadly rounded apex; anterior and posterior surfaces decline steeply; weakly translucent in profile ( Figures 6A View FIGURE 6 , 15–16 View FIGURE 15 View FIGURE 16 ).
Measurements. holotype ( n = 64). HL 1.63 (1.16–1.81); HW 1.66 (1.02–1.94); MOD 0.30 (0.21–0.34); OMD 0.39 (0.24–0.49); SL 1.25 (0.87–1.37); PNW 0.98 (0.69–1.19); HFL 1.51 (1.06–1.73); ML 2.01 (1.43–2.16); PW 0.30 (0.17–0.36); PPW 0.43 (0.27–0.52). Indices: SI 75.30 (67.03–94.58); CI 101.84 (80.56–119.5); OI 18.07 (16.4–22.66); HFI 90.96 (75.46–117.24).
Queen diagnosis. This caste is diagnosed by the following combination of features: (1) head and mesosoma dark orangish to orangish-brown with parts of mesopleura and mesoscutum blackish-orange to blackish; gaster blackishorange, (2) medial lobe of clypeus with strong submedial and sublateral rugae, (3) mandibles with 8 teeth, (4) dorsal base of scape weakly flattened; maximum basal width of scape greater than maximum preapical width, (5) MOD less than to greater than OMD, (6) cephalic dorsum weakly shining between fine, wavy to irregular, longitudinal rugae; medial rugae diverging toward posterior corners, rugae becoming strongly irregular to weakly rugoreticulate along posterior margin, (7) psammophore poorly developed, (8) sides of pronotum weakly shining between fine, longitudinal rugae; mesoscutum with fine, longitudinal rugae; mesoscutellum smooth and shining; anepisternum weakly shining between fine longitudinal rugae; katepisternum moderately shining between slightly more widely spaced rugae, (9) sides of propodeum weakly shining and densely punctate-granulate between longitudinal and oblique rugae; propodeal spines triangular, acuminate, length less than distance between their bases; infraspinal facet and propodeal declivity with fine transverse rugae, interrugae weakly dull to weakly shining, and (10) metasternal process large, higher than long with broadly rounded apex; anterior and posterior surfaces decline steeply; weakly translucent in profile ( Figure 17 View FIGURE 17 ).
Measurements. ( n = 5). HL 1.44–1.61; HW 1.39–1.58; MOD 0.32–0.35; OMD 0.28–0.36; SL 1.08–1.12; HFL 1.34–1.50; ML 2.10–2.26; PW 0.35–0.39; PPW 0.49–0.55. Indices: SI 68.35–79.14; CI 94.74–98.14; OI 22.22– 26.58; HFI 94.44–100.00.
Male diagnosis. This caste is diagnosed by the following combination of features: (11) head and mesosoma dark brown to dark blackish-brown; gaster a lighter medium brown, (12) clypeus weakly convex in profile; anteromedial margin weakly concave, with a weak medial longitudinal carina, (13) mandibles with 1 – 3 teeth basad of preapical tooth, (14) anterior ocellus well above level of top of eyes, (15) mesopleura weakly shining, moderately to strongly lineopunctulate-lineogranulate, sometimes with rugae along posterior margin,(16)propodeum contiguously punctategranulate and with scattered, irregular, fine rugae; propodeal spines cariniform to knoblike and inconspicuous, (17) metasternal process prominent and acute to elongate and narrowly rounded, and (18) subpetiolar process short, elongate-triangular to spiniform, apex acuminate to rounded ( Figures 1C View FIGURE 1 , 18 View FIGURE 18 ).
Measurements. ( n = 10). HL 0.84–1.01; HW 0.80–1.00; MOD 0.34–0.40; OMD 0.09–0.13; SL 0.34–0.49; HFL 1.43–1.90; ML 2.12–2.59; PW 0.28–0.39; PPW 0.46–0.55; AOD 0.09–0.13; IOD 0.26–0.34; OOD 0.26–0.35. Indices: SI 40.70–61.25; CI 80.00–114.94; OI 34.00–46.25; HFI 159.00–235.00.
Additional material examined. UNITED STATES: California: Butte Co.: Chico, 300’, May 18–19, 1956 & Aug 23, 1968 & no date (LACM; USNM); Neal Rd at 6.1 mi NE Jct Hwy 99, 1000’, Jun 23, 1979 (CASC; LACM); Hwy 99 at 15 mi S Chico, Apr 28, 1967 (LACM; UAIC; USNM); 9 km N Oroville, 360 m, Apr 18, 1992 (UCDC). Colusa Co.: Fouts Springs, 500 m, Apr 12, 1986 & May 28, 1995 (UCDC); Stonyford, 355 m, Apr 12, 1986 & Jun 19, 1988 (UCDC). Napa Co.: Twin Peaks at 4 km W Aetna Springs, 660 m, May 28, 1988 (UCDC). Sonoma Co.: 6 km N Sonoma, 300 m, Apr 13, 1997 (UCDC). Sutter Co.: 1 km W North Butte, Sutter Buttes, 200 m, May 28, 2007 (UCDC). Tehama Co.: Hwy 36 at 5.8 Rd mi E Jct Hwy 99, 550’, Jun 22, 1979 (CASC; LACM; USNM); Vina Plains Nature Preserve at 16 km E Corning, 60 m, Oct 10, 1992 & Mar 28, 1993 (UCDC); Dye Creek at 18 km SE Red Bluff, 335 m, Jun 30–Jul 2, 2002 (CASC). Tulare Co.: Woodlake (on label as “Wood L.”), May 14, 1947 (LACM); James K. Herbert Wetland Prairie Preserve, 310’, no date, 2004 (not examined, see Hamm & Kamansky, 2009) & Oct 2, 2018 (LACM; NHMW; RAJC; UCDC); Pixley National Wildlife Refuge, 200’, no date, 2004 (not examined, see Hamm & Kamansky, 2009) ( Figure 10C View FIGURE 10 ).
Etymology. This species was named based on the type locality at Chico, California.
Discussion. The poorly developed psammophore and short propodeal spines distinguish workers of V. chicoensis from all congeners except the closely related V. stoddardi . Veromessor chicoensis is distinguished by: (1) antennal scape narrowest near mid-length; maximum basal width of scape greater than maximum preapical width, and (2) in dorsal view, rugae on mesonotum similar to those on pronotum, interrugae weakly dull. In V. stoddardi : (1) antennal scape narrowest immediately distad of basal bend; maximum basal width of scape about equal to maximum preapical width, and (2) in dorsal view, the mesonotum usually smooth and shining, rugae distinctly weaker than those on pronotum.
Smith (1956) described V. stoddardi chicoensis from a holotype and 20 paratype workers and indicated specimens were deposited at USNM. We examined nine type workers (on three pins) from USNM. All nine had type labels USNM #62959, but none of these specimens had a holotype label. Two pins had a USNM Paratype No. 62959 label, and one pin had a USNM Type No. 62959 label; the latter pin also had a black dot on the uppermost triangle, and this specimen is presumed to be the holotype. Consequently, we placed a holotype label on this worker.
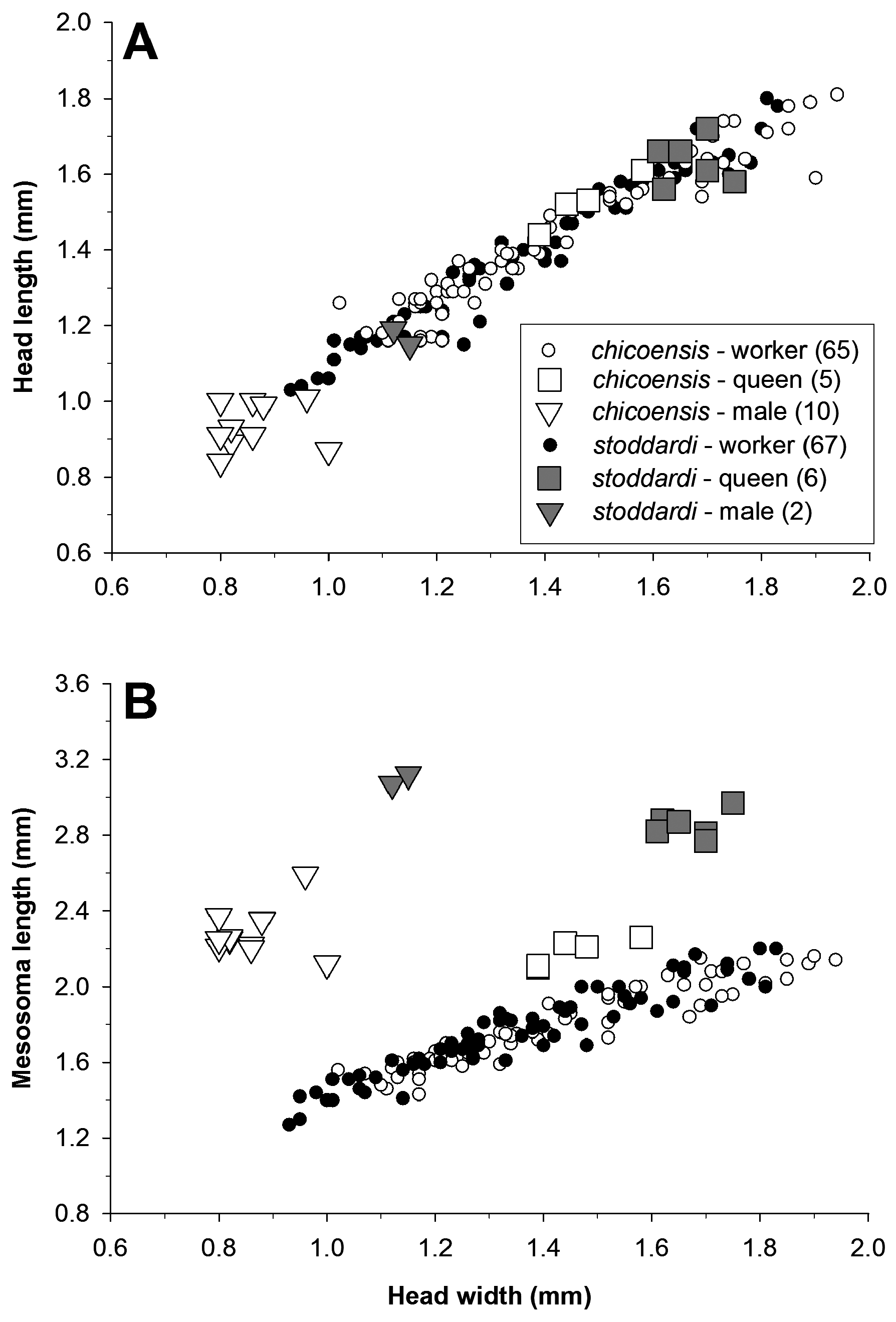
Veromessor chicoensis was described as a subspecies of V. stoddardi because of the similarity between workers of the two species. The two species are allopatric and workers of the two species display few consistent morphological differences, but queens and males differ in both morphology and size. Worker allometry is similar for both species (see above), but queens and males of V. chicoensis are distinctly smaller than those of V. stoddardi ( Figure 19 View FIGURE 19 ). A molecular phylogeny that used UCEs shows V. chicoensis and V. stoddardi to be sister lineages, but well-separated and reciprocally monophyletic (M.L. Borowiec, unpub. data).
As discussed by Hamm and Kamansky (2009), workers from an apparently disjunct population in Tulare County, California (see Figure 10C View FIGURE 10 and below), differed slightly from those in northern populations with rugae on the head and mesosoma less pronounced with the interrugae more shining. A molecular phylogeny based on UCEs shows that these populations show little divergence compared to interspecific genetic distances in the genus (M.L. Borowiec, unpub. data). Specimens referred to in Hamm and Kamansky (2009) were deposited in LACM, but could not be located, and they are presumed lost.
Biology. Little is known about the biology of V. chicocensis . Workers are diurnal foragers during spring and summer months, but shift to crepuscular foraging at temperatures of about 30 o C and forage nocturnally at temperatures of 35 o C ( Hamm & Kamansky, 2009). Previous indications were that V. chicoensis workers were solitary foragers based on “no conspicuous foraging trail was observed” (M. Bennett, pers. comm.). However, the senior author observed two colonies in Tulare County in early October 2018 that were foraging in a narrow 5–10 m long column during mid-morning. Colonies appeared to contain about 1,000 –2,000 workers (R.A. Johnson, pers. obs). Workers are polymorphic ( Figure 19 View FIGURE 19 ). Nests are placed in hard clay soils. Veromessor chicoensis is sometimes sympatric with V. andrei , whereas it appears to occur without V. andrei at other locations, especially those with marginal, thin soil habitats (P.S. Ward, pers. comm.).
Mating flights have not been observed, but sexuals have been collected from May 14 through August 28, suggesting that mating flights start early in the summer and last for an extended period. Nothing is known about colony founding, but head width and head length of queens of V. chicoensis are about the same as medium-sized workers, whereas mesosoma length is greater in queens than in workers ( Figure 19 View FIGURE 19 ). Their small size infers that founding queens of V. chicoensis are semiclaustral, probably obligate foragers.
Veromessor chicoensis is also unusual because dealate queens sometimes occur in foraging columns (one foraging dealate queen collected by the senior author on October 2, 2018). Foraging by dealate queens is common in V. andrei , but this is the first such record in V. chicoensis (see above discussion under V. andrei ).
Gland chemistry has not been examined in V. chicoensis . Like other small-colony congeners, workers of V. chicoensis have a small pygidial gland reservoir and lack a textured tergal cuticle ( Hölldobler et al., 2013).
The Pacific Coast horned lizard ( Phrynosoma cornutum ) preys upon V. chicoensis , and it sometimes is the main component in the diet of these lizards ( Hamm & Kamansky, 2009).
Veromessor chicoensis appears to be restricted to the Sacramento and San Joaquin Valleys in northern and central California and in low mountains along the northwestern edge of the Sacramento Valley. Veromessor chicoensis probably occurred throughout most to all of the San Joaquin Valley historically given a 1947 record of one male from Tulare County, and more recent collections from Tulare County near vernal pools in Pixley National Wildlife Refuge and James K. Herbert Wetland Prairie Preserve ( Hamm & Kamansky, 2009). The senior author collected two colonies in the James K. Herbert Wetland Prairie Preserve in October 2018, but this species was not located during a brief survey at Pixley National Wildlife Refuge at that time. Moreover, the current range of V. chicoensis in the San Joaquin Valley appears to be limited to isolated properties and refuges that have not been developed for agriculture ( Hamm & Kamansky, 2009).
This species occurs at elevations from 60–660 m in the California interior chaparral and woodlands and California Central Valley grasslands ecoregions, as defined by Olson et al. (2001) ( Figure 10C View FIGURE 10 ).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Myrmicinae |
|
Genus |
Veromessor chicoensis
| Johnson, Robert A., Borowiec, Marek L., Snelling, Roy R. & Cole, Arthur C. 2022 |
Veromessor stoddardi subsp. chicoensis
| Smith, M. R. 1956: 37 |