
Chamberlainium capense Puckree-Padua, P.W. Gabrielson et Maneveldt, 2021
|
publication ID |
https://doi.org/ 10.1515/bot-2020-0074 |
|
persistent identifier |
https://treatment.plazi.org/id/039387E3-FF8C-FF95-8838-0621071BFFE5 |
|
treatment provided by |
Felipe |
|
scientific name |
Chamberlainium capense Puckree-Padua, P.W. Gabrielson et Maneveldt |
| status |
sp. nov. |
Chamberlainium capense Puckree-Padua, P.W. Gabrielson et Maneveldt sp. nov. ( Figures 4 View Figures 4–6 , 7–11 View Figures 7–11 , 12–14 View Figures 12–14 , 15–18 View Figures 15–18 ; Tables 1, 2).
Holotype: L 3986119 , 09.x.2016, leg. G. W. Maneveldt, collection number 16/05, epilithic on primary bedrock in mid-intertidal, sand inundated rock pool.
Type locality: South Africa, Western Cape Province, Mouille Point (33°53.9440′ S, 18°24.5285′ E) GoogleMaps .
Etymology: ‘ capense ’ in reference to the species’ restricted distribution along the Cape Peninsula historically known as the ‘ Cape of Good Hope, South Africa’ (Caput Bonae Spei).
Description: Non-geniculate, thalli are moderately thick (up to 1000 µm), encrusting to mostly variably lumpy to slightly protuberant. Thalli are epilithic or epizoic and bright to dusky pink in well-lit conditions. Individual crusts do not appear to fuse together and are easily discernible. Thallus construction is monomerous with a single layer of epithallial cells. A central columella is present in tetrasporangial conceptacles that disintegrate to form a low mound with maturity. The pore opening in mature tetrasporangial conceptacles is occluded by a corona of filaments that projects above the opening. The psb A (851 bp) and rbc L (691–1387 bp) sequences are diagnostic.
Habitat: Thalli were epilithic on the primary bedrock and on large boulders in rock pools and on exposed platforms in the mid-intertidal zone, as well as epizoic on mollusc shells in the low intertidal zone.
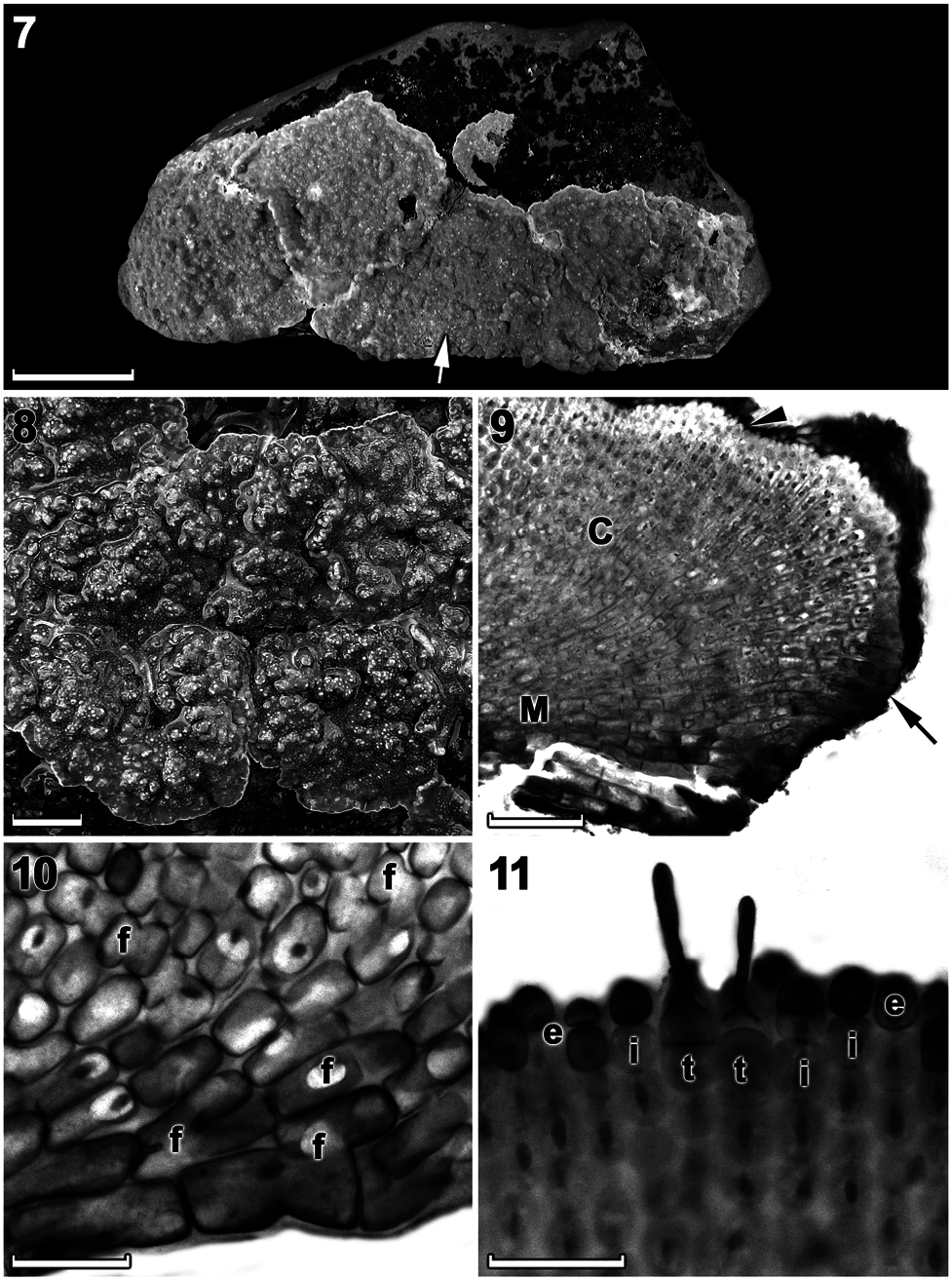
Vegetative morphology and anatomy: Thalli were non-geniculate, moderately thick (up to 1000 μ m), encrusting (smooth) to mostly variably lumpy and slightly protuberant, with protuberances to 4 mm tall ( Figures 4 View Figures 4–6 and 7, 8 View Figures 7–11 ), and were firmly adherent, dusky pink to mauve (in well-lit conditions) to rosy or purple-pink (in dim light) when freshly collected ( Figure 4 View Figures 4–6 ). Individual crusts did not fuse together and were easily discernible ( Figures 7, 8 View Figures 7–11 ).
Thalli were dorsiventrally organized, monomerous and haustoria were absent. The medulla was thin and plumose (non-coaxial) ( Figures 9, 10 View Figures 7–11 ). Medullary filaments comprised rectangular to elongate cells, which gave rise to cortical filaments that comprised mainly square to rectangular cells ( Figure 10 View Figures 7–11 ). Contiguous medullary and cortical filaments were joined by cell fusions; secondary pit connections were absent ( Figure 10 View Figures 7–11 ). Subepithallial initials (intercalary meristematic cells) were square to rectangular ( Figure 11 View Figures 7–11 ). The epithallus was single layered with oval to rounded cells ( Figure 11 View Figures 7–11 ). Trichocytes were common at the thallus surface and occurred singularly (mostly) to paired ( Figure 11 View Figures 7–11 ). Trichocytes were always terminal and never intercalary in the cortex; buried trichocytes were not observed. Data on morphological and measured vegetative characters are summarized in Table 1.
Reproductive morphology and anatomy: Gametangial thalli appeared to be dioecious, although female plants were not observed.
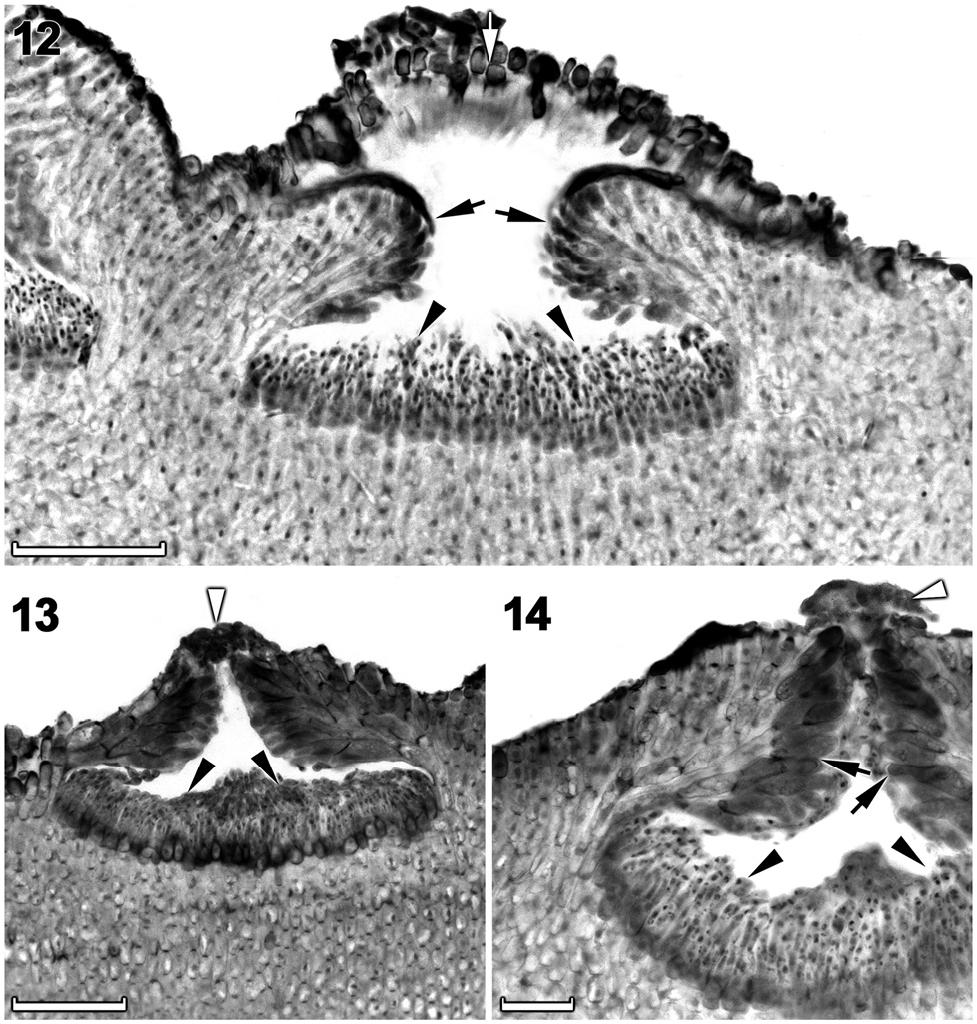
Spermatangial (male) conceptacles were uniporate, low-domed and raised above surrounding thallus surface ( Figures 12–14 View Figures 12–14 ). Conceptacle chambers were transversely elliptical to flatten; conceptacle roof nearly twice as thick along pore canal ( Figures 13, 14 View Figures 12–14 ). Conceptacle roof formed from filaments peripheral to the fertile area ( Figure 12 View Figures 12–14 ). Throughout the early development, a protective layer of epithallial cells surrounded the conceptacle primordium ( Figure 12 View Figures 12–14 ). This protective layer was shed once the pore canal was near fully developed. The pore opening was occluded by a mucilage plug ( Figures 13, 14 View Figures 12–14 ). In mature conceptacles, terminal initials along pore canal were enlarged and papillate; they projected into the pore canal and were orientated more or less parallel to the conceptacle roof surface ( Figure 14 View Figures 12–14 ). Unbranched (simple) spermatangial systems were confined to mature conceptacle floor ( Figures 12–14 View Figures 12–14 ). Senescent male conceptacles appeared to be shed; no buried conceptacles were observed.
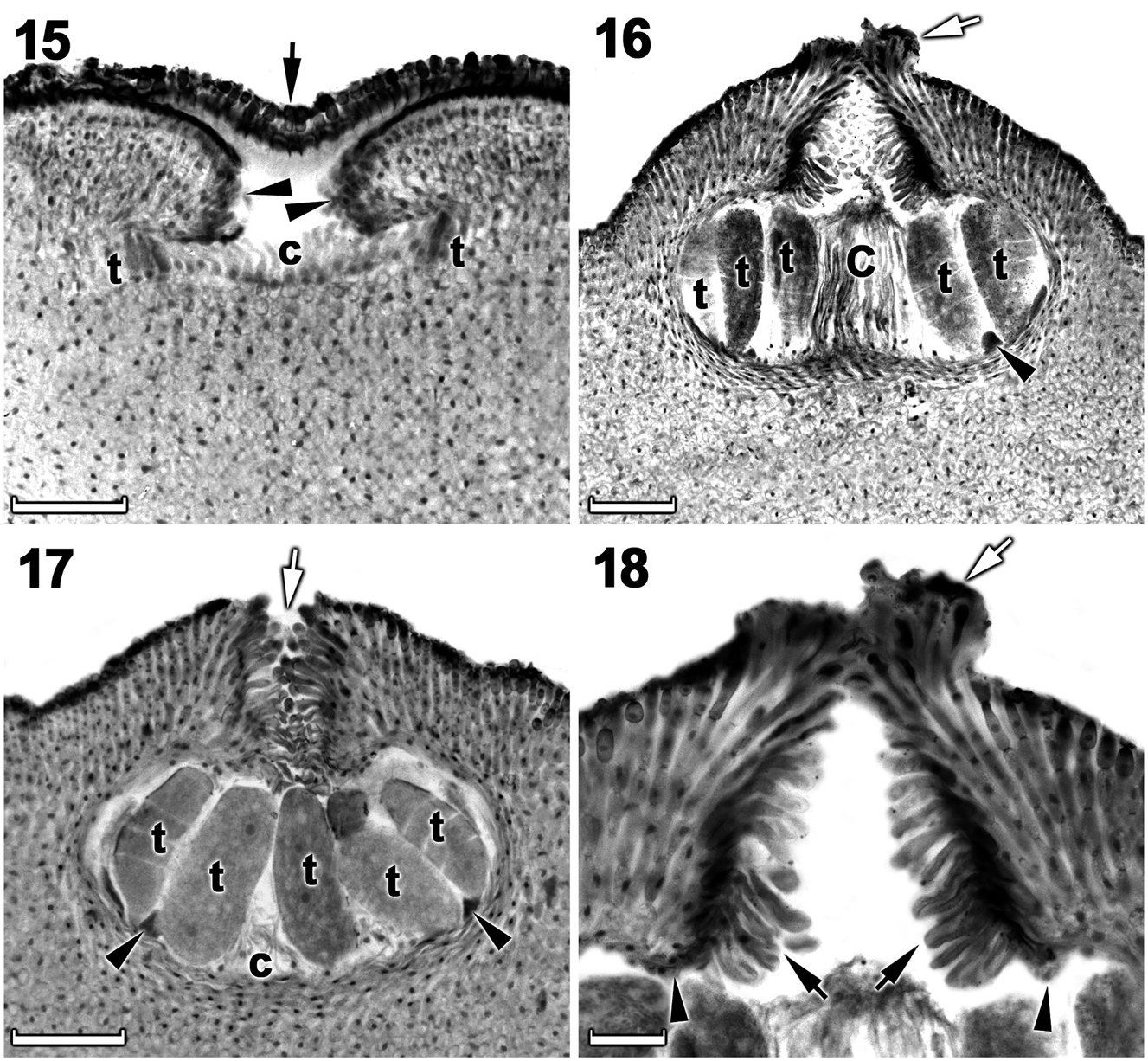
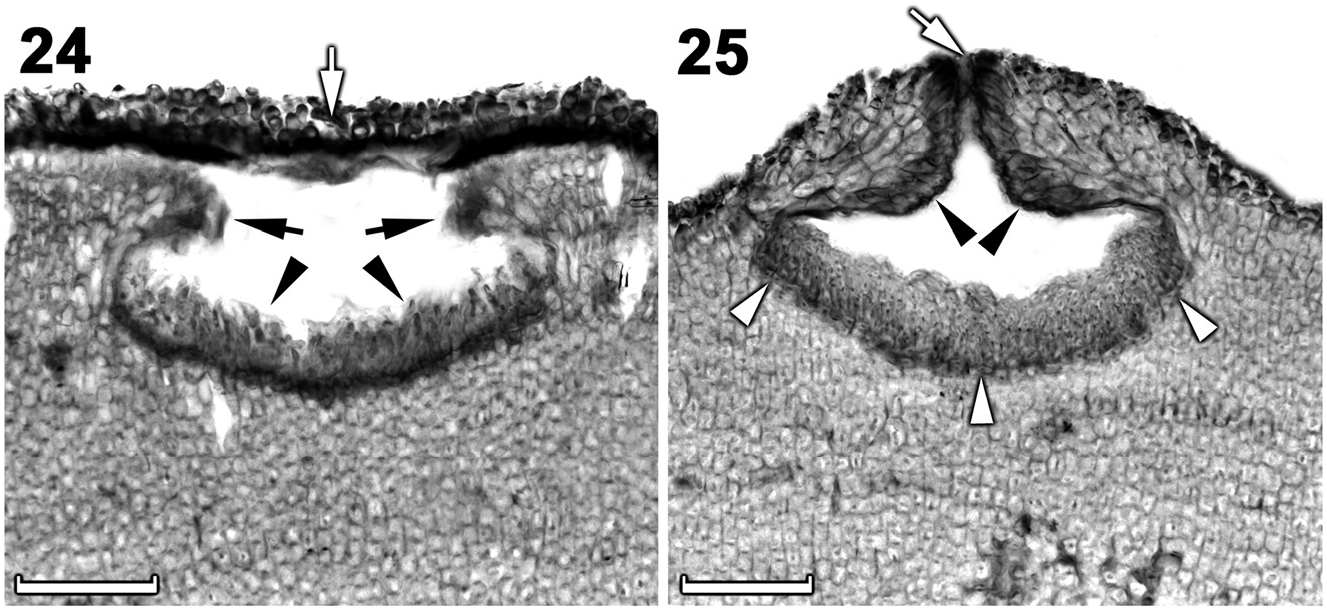
Tetrasporangial thalli were morphologically similar to spermatangial thalli. Conceptacles were uniporate, low domed and raised above the surrounding thallus surface ( Figures 15–17 View Figures 15–18 ). Conceptacle chambers were transversely elliptical to bean-shaped. Conceptacle roof was nearly twice as thick along the pore canal and was 5–7 cells thick, including an epithallial cell. Pore canal tapered towards the surface, formed an arch ( Figure 18 View Figures 15–18 ) and was lined by elongated papillate cells that projected into the canal and were orientated more or less parallel or nearly perpendicular to the conceptacle roof surface ( Figure 18 View Figures 15–18 ). The roof was formed from filaments peripheral to the fertile area ( Figure 15 View Figures 15–18 ) and terminal initials were more elongate than their inward derivatives. Throughout early development a protective layer of epithallial cells surrounded the conceptacle primordium ( Figure 15 View Figures 15–18 ). This protective layer was shed once pore canal was near fully developed; the pore opening was occluded by a corona of filaments that projected above the pore opening ( Figures 16, 18 View Figures 15–18 ). The corona appeared to form from filaments near the upper half of the roof, directly adjacent to the pore canal ( Figures 16, 18 View Figures 15–18 ). Throughout the development of the tetrasporangial conceptacle, a prominent columella of sterile filaments formed at the center of the conceptacle chamber ( Figure 15 View Figures 15–18 ), which extended into the pore canal ( Figure 16 View Figures 15–18 ); the central columella appeared weakly calcified as with maturity it disintegrated to form a low mound ( Figure 17 View Figures 15–18 ). The base of the pore canal was sunken into the chamber and terminal initials near the base pointed downward ( Figure 18 View Figures 15–18 ). Mature conceptacle floors were sunken 11– 17 cells (including the epithallial cell) below the surrounding thallus surface. Zonately divided tetrasporangia were arranged at the extreme periphery of the conceptacle chamber and were attached via a stalk cell ( Figures 16, 17 View Figures 15–18 ). Where the central columella had disintegrated or diminished, the tetrasporangia filled the chamber and appeared to be distributed across the chamber floor ( Figure 17 View Figures 15–18 ). Senescent tetrasporangial conceptacles appeare to be shed as no buried conceptacles were observed. Data on reproductive characters are summarized in Table 2.
Distribution: Confirmed by DNA sequences to have a restricted distribution (± 43 km distance) along the southwest coast, from Mouille Point to Slangkop (Western Cape Province) along the Cape Peninsula, South Africa.
Chamberlainium glebosum Puckree-Padua, P.W. Gabrielson et Maneveldt sp. nov. ( Figures 5 View Figures 4–6 , 19–23 View Figures 19–23 , 24–25 View Figures 24–25 , 26–30 View Figures 26–30 ; Tables 1, 2).
Holotype: L 3986123 , 17vii.2015, leg. G. W. Maneveldt, collection number 18/05, epilithic on primary bedrock in a mid-intertidal rock pool.
Type locality: South Africa, Western Cape Province, Melkbosstrand (33°44.2518′ S, 18°26.1343′ E) GoogleMaps .
Etymology: ‘ glebosum ’ from ‘ glebosus ’, making reference to the species’ lumpy ( Stearn 1973) and highly protuberant growth form.
Description: Non-geniculate, thalli are thick (to 2000 µm), very lumpy becoming highly protuberant. Thalli are epilithic and brownish pink to gray in well-lit conditions. Individual crusts do not appear to coalesce (do not fuse together) and are easily discernible. The thallus construction is monomerous with a single layer of epithallial cells. A central columella is present in tetrasporangial conceptacles, which disintegrates to form a low mound with maturity. The pore opening in mature tetrasporangial conceptacles is unoccluded. The psb A (851 bp) and rbc L (691–1387 bp) gene sequences are diagnostic.
Habitat: Thalli were epilithic on the primary bedrock in the high and mid-intertidal zones and only occasionally on the low shore.
Vegetative morphology and anatomy: Thalli were non-geniculate, thick (up to 2000 μ m), very lumpy
Character Chamberlainium Chamberlainium Chamberlainium cochle- Chamberlainium Chamberlainium Chamberlainium nata- Chamberlainium occiagulhense (van der capemse (this study) are (Puckree-Padua et al. glebosum (this impar (Puckree- lense (Puckree-Padua dentale (this study) Merwe et al. zeIs) zezeb) study) Padua et al. zezeb) et al. zezeb)
Growth form Encrusting (smooth) Encrusting (smooth), Encrusting (smooth) Lumpy, becoming Encrusting (smooth), Encrusting (smooth) Encrusting (smooth) to becoming variably highly protuberant orbicular becoming warty (mostly) to lumpy, lumpy (mostly) and con fl uent becoming slightly slightly protuberant protuberant
Thallus Dimerous Monomerous Monomerous Monomerous Monomerous Monomerous Monomerous construction
Maximum thallus 650 1000 370 2000 1800 800 1000
thickness
Habit Individual thalli Individual thalli Individual thalli not Individual thalli Individual thalli Individual thalli discern- Individual thalli discerndiscernible, not discernible, not fusing discernible, fusing to discernible, not discernible, not ible, not fusing ible, not fusing
fusing form large expanses fusing fusing
Trichocytes None observed Common, solitary Common, solitary None observed Rare, solitary to Rare, solitary (mostly) to Common, solitary (mostly) to paired (mostly) to clustered, up paired clustered, up to 6 (sepa- (mostly) to clustered, up to 6 (separated by normal rated by normal vegeta- to 6 (separated by normal vegetative fi laments) tive fi laments) vegetative fi laments) Trichocyte - L = 22–29; D = 5–7 L = 15–27; D = 7–12 - L = 22–39; D = 10– L = 12–29; D = 7–10 L = 22–37; D = 7–10 dimensions 12
No. of epithallial 1–3 (mostly 1–2) 1 1 1 4–6 (9) 1 1
cell layers
Epithallial cell Rounded to domed Oval to round Elliptical to domed Rounded to Elliptical to rounded Flattened to elliptical Elliptical to rounded to shape elliptical elongate
Epithallial cell ND L = 5–7; D = 5–7 L = 2–7; D = 5–7 L = 5–7; D = 2–7 L = 5–7; D = 5–7 L = 5–7; D = 5–7 L = 5–8; D = 2–7 dimensions
Subepithallial cell Square to rectangular Square to rectangular Square to rectangular Rectangular Square to Square to rectangular Rectangular
shape rectangular
Subepithallial cell ND L = 5–15; D = 5–10 L = 3–10; D = 4–7 L = 5–12; D = 2–7 L = 5–12; D = 5–7 L = 7–10; D = 5–7 L = 10–12; D = 5–7 dimensions
Cortical cell shape - Square to rectangular Square to rectangular Square to Square to beaded Square to rectangular Square to rectangular rectangular
Cortical cell - L = 7–10; D = 7–10 L = 5–12; D = 4–12 L = 5–15; D = 5–0 L = 7–10; D = 2–5 L = 7–25; D = 5–7 L = 7–10; D = 7–12 dimensions
Medullary cell - Rectangular Rectangular to elongate Rectangular to Rectangular to Rectangular to elongate Square to rectangular shape elongate elongate
Medullary cell - L = 22–44; D = 15–25 L = 10–30; D = 3–16 L = 12–25; D = 7– L = 5–25; D = 7–14 L = 15–20 D = 5–7 L = 15–17; D = 7–10 dimensions 12
Erect fi lament cell Square to rectangular - - - - - -
shape with rounded corners
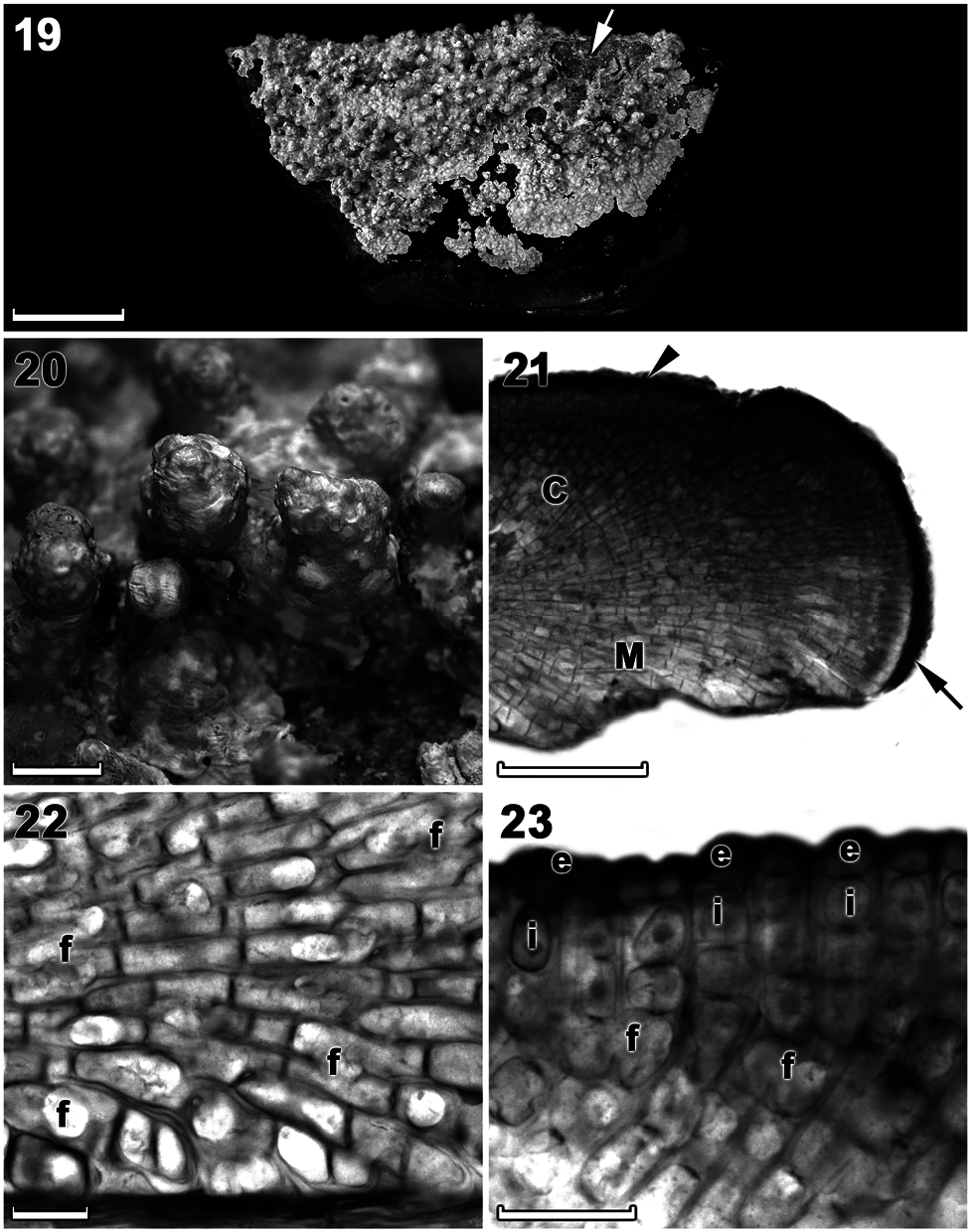
becoming highly protuberant with protuberances to 6 mm tall ( Figures 5 View Figures 4–6 , 19 and 20 View Figures 19–23 ), and were firmly adherent, mostly brownish pink to greyish when freshly collected ( Figure 5 View Figures 4–6 ). Individual crusts did not coalesce (did not fuse together) and were easily discernible ( Figure 5 View Figures 4–6 ).
Thalli were dorsiventrally organized, monomerous and haustoria were absent. The medulla was thin and plumose (non-coaxial) ( Figures 21, 22 View Figures 19–23 ). Medullary filaments comprised rectangular to elongate cells, which gave rise to cortical filaments that comprised square to rectangular cells ( Figures 22, 23 View Figures 19–23 ). Contiguous medullary and cortical filaments were joined by cell fusions; secondary pit connections were absent ( Figures 22, 23 View Figures 19–23 ). Subepithallial initials (intercalary meristematic cells) were square to rectangular ( Figure 23 View Figures 19–23 ). The epithallus was single layered with rounded to elliptical cells ( Figure 23 View Figures 19–23 ). Trichocytes were not observed. Data on morphological and measured vegetative characters are summarized in Table 1.
Reproductive morphology and anatomy: Gametangial thalli appeared to be dioecious, although female plants were not observed.
Spermatangial (male) conceptacles were uniporate, low-domed, raised above surrounding thallus surface ( Figures 24, 25 View Figures 24–25 ). Conceptacle chambers were transversely elliptical to flatten with the roof nearly twice as thick along the pore canal ( Figure 25 View Figures 24–25 ). The roof was formed from filaments peripheral to the fertile area ( Figure 24 View Figures 24–25 ). Throughout the early development, a protective layer of epithallial cells surrounded the conceptacle primordium ( Figure 24 View Figures 24–25 ). This protective layer was shed once the pore canal was near fully developed. The pore opening was occluded by a mucilage plug ( Figure 25 View Figures 24–25 ). In mature conceptacles the terminal initials along the pore canal were enlarged and papillate, they projected into the pore canal and were orientated more or less parallel to the conceptacle roof surface ( Figure 25 View Figures 24–25 ). Unbranched (simple) spermatangial systems were confined to floor of mature conceptacle ( Figures 24, 25 View Figures 24–25 ). Senescent male conceptacles appeared to be shed as no buried conceptacles were observed.
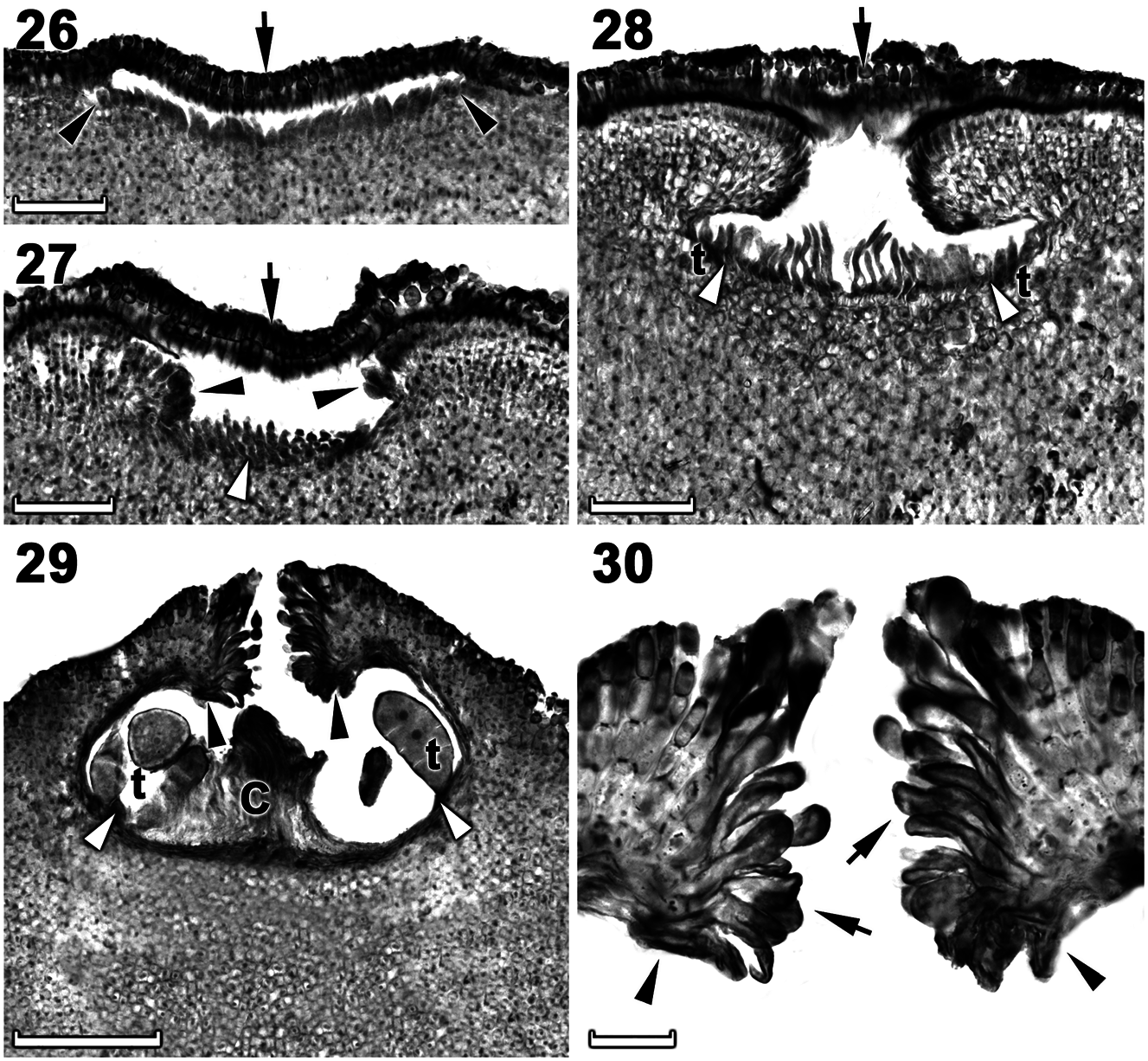
Tetrasporangial thalli were morphologically similar to spermatangial thalli. Conceptacles were uniporate, low domed and raised above surrounding the thallus surface ( Figures 26–29 View Figures 26–30 ). Conceptacle chambers were transversely elliptical to bean-shaped. The roof was nearly twice as thick along the pore canal and were 4–8 (9) cells (including an epithallial cell) thick ( Figure 29 View Figures 26–30 ). The pore canal tapered towards the surface was lined by elongated papillate cells that projected into the pore canal and were orientated more or less parallel or nearly perpendicular to the conceptacle roof surface ( Figure 30 View Figures 26–30 ). The roof was formed from filaments peripheral to the fertile area ( Figures 26–28 View Figures 26–30 ) and terminal initials were more elongate than their inward derivatives ( Figure 27 View Figures 26–30 ). Throughout the early development a protective layer of epithallial cells surrounded the conceptacle primordium ( Figures 26–28 View Figures 26–30 ). This protective layer was shed once the pore canal was near fully developed. The pore opening in mature conceptacles was unoccluded ( Figures 29, 30 View Figures 26–30 ). Throughout the development of the immature tetrasporangial conceptacle, a prominent columella of sterile filaments formed at the center of the conceptacle chamber ( Figures 27, 28 View Figures 26–30 ), which extended into the pore canal ( Figure 29 View Figures 26–30 ); the central columella appeared weakly calcified as with maturity it disintegrated to form a
low mound. The base of the pore canal was sunken into the chamber and terminal initials near the base pointed downward ( Figures 29, 30 View Figures 26–30 ). Mature conceptacle floors were sunken 11–24 cells (including the epithallial cell) below the surrounding thallus surface. Zonately divided tetrasporangia at maturity were arranged at the extreme periphery of conceptacle chamber and was attached via a stalk cell ( Figure 29 View Figures 26–30 ). Senescent tetrasporangial conceptacles appeared to be shed as no buried conceptacles were observed. Data on reproductive characters are summarized in Table 2.
Distribution: Confirmed by DNA sequences to have a wide, but disjunct distribution along the west coast, occurring from Port Nolloth to Groenriviermond (Northern Cape Province, ± 220 km distance) and then from Cape St. Martin to Slangkop (Western Cape Province, ± 200 km distance), South Africa.
Chamberlainium occidentale Puckree-Padua, P.W. Gabrielson et Maneveldt sp. nov. ( Figures 6 View Figures 4–6 , 31–35, 36–41 View Figures 36–41 , 42–46 View Figures 42–46 ; Tables 1, 2).
Holotype: L 3986120 , 01.x.2015, leg. G. W. Maneveldt and C. A. Puckree-Padua, collection number 15/61 B, epilithic on primary bedrock in low intertidal zone.
Isotypes: UWC 15 View Materials /58, UWC 15 View Materials /59, UWC 15 View Materials /61 A
Type locality: South Africa, Western Cape Province, Lamberts Bay (32°6.5728′ S, 18°18.1270′ E) GoogleMaps .
Etymology: ‘ occidentale ’ from ‘ occidentalis ’, meaning western ( Stearn 1973), making reference to the species’ widespread distribution along the west coast of Southern Africa.
Description: Non-geniculate, thalli are variably thin to thick (up to 1000 µm), encrusting (smooth) to warty (mostly) to lumpy, becoming only slightly protuberant. Thalli are epilithic to epizoic and brownish beige in well-lit conditions. Individual crusts do not appear to coalesce (do
not fuse together) and are easily discernible. The thallus construction is monomerous with a single layer of epithallial cells. A central columella is present in tetrasporangial conceptacles, which persists to maturity. The pore opening in mature tetrasporangial conceptacles is unoccluded and slightly sunken below the surrounding roof surface. The psb A (851 bp) and rbc L (691–1,384 bp) sequences are diagnostic.
Habitat: Thalli were epilithic on the primary bedrock in the high, mid- and low intertidal zones and epizoic on mollusc shells in the low intertidal zone.
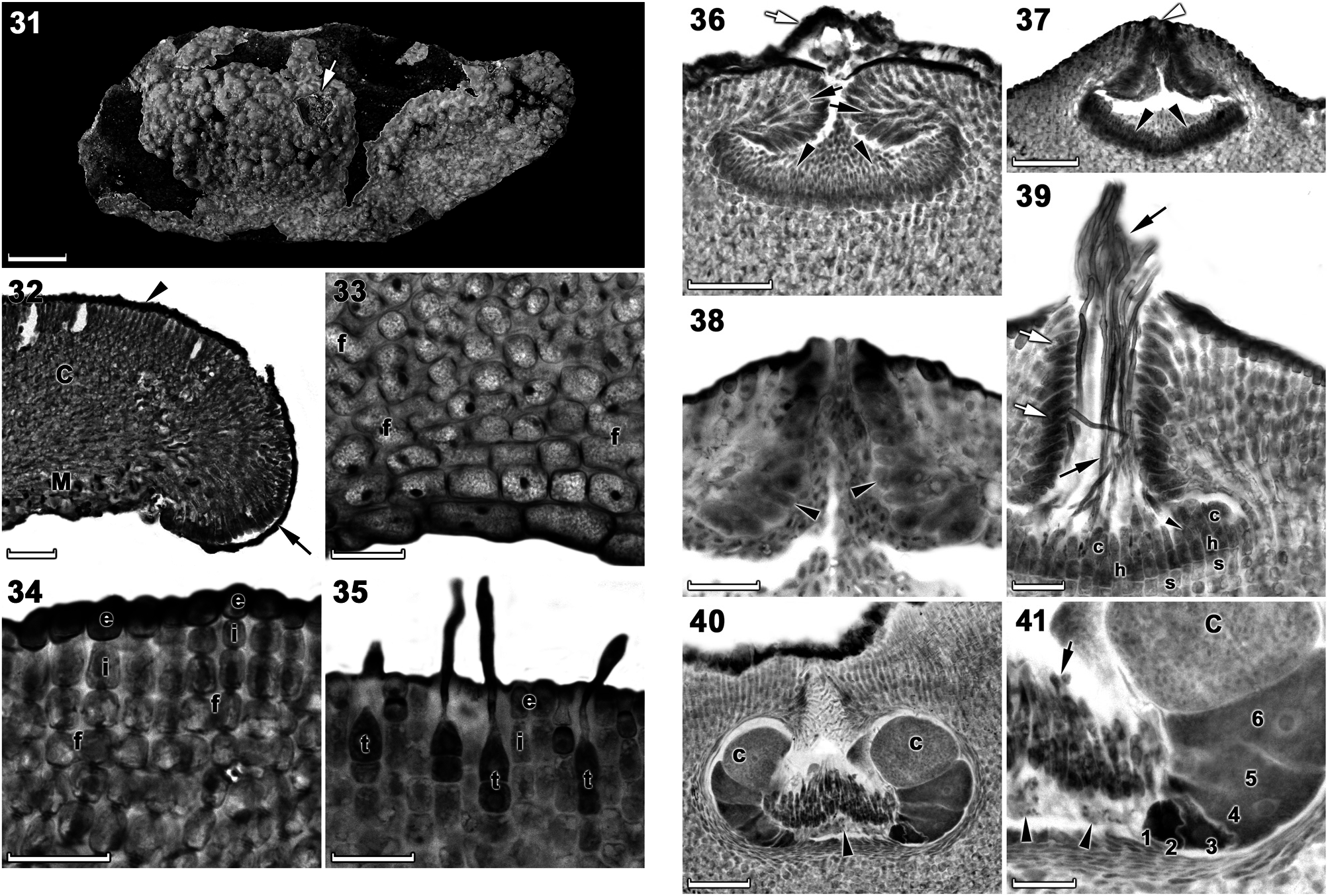
Vegetative morphology and anatomy: Plants were non-geniculate, thalli were variably thin to thick (up to 1000 μ m), encrusting (smooth) to warty (mostly) to lumpy, becoming only slightly protuberant (protuberances 4 mm in height) ( Figures 6 View Figures 4–6 , 31). Thalli were firmly adherent, brownish beige (in well-lit conditions) ( Figures 6 View Figures 4–6 ) to bluegray to rosy pink (in dim light) when freshly collected. Individual crusts did not coalesce (did not fuse together) and were easily discernible ( Figure 31).
Thalli were dorsiventrally organized, monomerous and haustoria were absent. The medulla was thin and plumose (non-coaxial) ( Figures 32, 33). Medullary filaments comprised square to elongate cells, which gave rise to cortical filaments that comprised square to rectangular cells ( Figures 33, 34). Contiguous medullary and cortical filaments were joined by cell fusions; secondary pit connections were absent ( Figures 33, 34). Subepithallial initials were square to rectangular ( Figure 35). The epithallus was single layered with elliptical to round to elongate cells ( Figure 35). Trichocytes were common, mostly singularly, but also in clusters of up to 6 cells, separated by normal vegetative filaments ( Figure 35). Trichocytes were always terminal and never intercalary in the cortex; buried trichocytes were not observed. Data on morphological and measured vegetative characters are summarized in Table 1.
Reproductive morphology and anatomy: Gametangial thalli were dioecious and monoecious. The type specimen bore all (including tetrasporangial) life cycle stages.
Spermatangial (male) conceptacles were uniporate, low-domed, and raised above surrounding thallus surface ( Figures 36, 37 View Figures 36–41 ). Conceptacle chambers were transversely elliptical to flatten and the roof nearly twice as thick along the pore canal ( Figures 37, 38 View Figures 36–41 ). The roof was formed from filaments peripheral to the fertile area ( Figure 36 View Figures 36–41 ). Throughout the early development, a protective layer of epithallial cells surrounded the conceptacle primordium ( Figure 36 View Figures 36–41 ). This protective layer was shed once the pore canal was near fully developed. The pore opening was occluded by a mucilage plug ( Figure 37 View Figures 36–41 ). In mature conceptacles terminal initials along the pore canal were enlarged and papillate, they projected into the pore canal and were orientated more or less parallel to the conceptacle roof surface ( Figure 38 View Figures 36–41 ). Unbranched (simple) spermatangial systems were confined to the floor of the mature conceptacle ( Figures 36, 37 View Figures 36–41 ). Senescent male conceptacles appeared to be shed as no buried conceptacles were observed.
Carpogonial (female) conceptacles were similar in size and shape to spermatangial conceptacles and were raised above surrounding thallus surface ( Figure 39 View Figures 36–41 ). In mature conceptacles the terminal initials along the pore canal were enlarged and papillate, they projected into the pore canal and were orientated more or less parallel or nearly perpendicular to the conceptacle roof surface ( Figure 39 View Figures 36–41 ). Carpogonial branches developed across the floor of the conceptacle chamber, comprising a single support cell, a hypogynous cell and a carpogonium that extended into a trichogyne ( Figure 39 View Figures 36–41 ). Sterile cells were occasionally present on hypogynous cells ( Figure 39 View Figures 36–41 ).
After presumed karyogamy, carposporophytes developed within female conceptacles and formed carposporangial conceptacles ( Figure 40 View Figures 36–41 ). Carposporangial conceptacles were comparatively large, low-domed, and raised above the surrounding thallus surface. Conceptacle chambers were transversely elliptical with flattened bottoms presumably caused by the growth of the gonimoblast filaments. Pore canals tapered towards the surface, were lined by enlarged papillate cells that projected into the pore canal and were orientated more or less parallel to the conceptacle roof surface ( Figure 40 View Figures 36–41 ). In mature conceptacles, the cells lining the pore canal were more elongate than their inward derivatives. The base of the pore canal was sunken into the chamber and terminal initials near the base characteristically pointed downward ( Figure 40 View Figures 36–41 ). A discontinuous central fusion cell was present and 5–7 celled gonimoblast filaments (including a terminal carposporangium) developed along the periphery of the conceptacle chamber ( Figures 40, 41 View Figures 36–41 ). The remains of unfertilized carpogonia persisted across the dorsal surface of the fusion cell ( Figures 40, 41 View Figures 36–41 ). Senescent carposporangial conceptacles appeared to be shed as no buried conceptacles were observed.
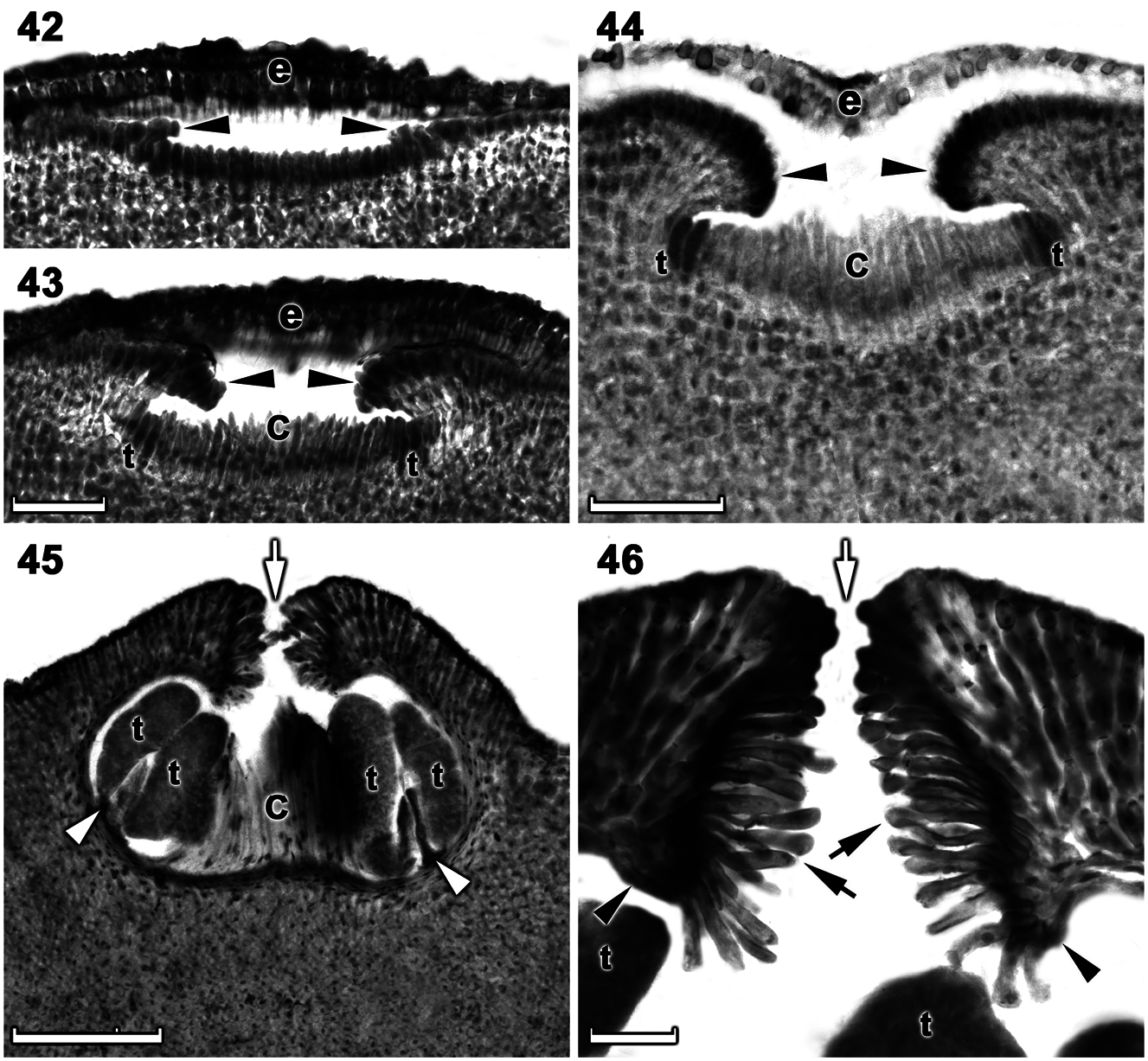
Tetrasporangial thalli are morphologically similar to gametangial thalli. Conceptacles were uniporate, low domed and raised above the surrounding thallus surface ( Figures 42–45 View Figures 42–46 ). Conceptacle chambers were transversely elliptical to bean-shaped. The roof was nearly twice as thick along the pore canal and was 5–7 cells (including an epithallial cell) thick. The pore canal tapered towards the surface an was arched ( Figure 46 View Figures 42–46 ). The pore canal was lined by elongated papillate cells that projected into the pore canal and were orientated more or less parallel or nearly perpendicular to the conceptacle roof surface ( Figure 46 View Figures 42–46 ). The roof was formed from filaments peripheral to the fertile area and terminal initials were more elongate than their inward derivatives ( Figures 42–44 View Figures 42–46 ). Throughout the early development a protective layer of epithallial cells surrounded the conceptacle primordium ( Figures 42–44 View Figures 42–46 ). This protective layer was shed once the pore canal was near fully developed. The pore opening was unoccluded and became slightly sunken below the surrounding roof surface ( Figures 45, 46 View Figures 42–46 ). Throughout development of the immature tetrasporangial conceptacle a prominent columella of sterile filaments formed at the center of the conceptacle chamber ( Figures 43, 44 View Figures 42–46 ), which persisted to maturity ( Figure 45 View Figures 42–46 ). The base of the pore canal was sunken into the chamber and terminal initials near the base point downward ( Figure 46 View Figures 42–46 ). Mature conceptacle floors were sunken 14–19 cells (including the epithallial cell) below the surrounding thallus surface. Zonately divided tetrasporangia at maturity were arranged at the extreme periphery of the conceptacle chamber and were attached via a stalk cell ( Figure 45 View Figures 42–46 ). Senescent tetrasporangial conceptacles appeared to be shed as no buried conceptacles were observed. Data on reproductive characters summarized in Table 2.
Distribution: Confirmed by DNA sequences to be widely distributed (± 1,200 km distance) along the west and southern west coasts, occurring from Lüderitz ( Namibia) to Holbaaipunt (Western Cape Province), South Africa. The species is disjunct in the southern part of its range of ± 240 km.

| G |
Conservatoire et Jardin botaniques de la Ville de Genève |
| W |
Naturhistorisches Museum Wien |
| A |
Harvard University - Arnold Arboretum |
| L |
Nationaal Herbarium Nederland, Leiden University branch |
| ND |
University of Notre Dame |
| C |
University of Copenhagen |
| B |
Botanischer Garten und Botanisches Museum Berlin-Dahlem, Zentraleinrichtung der Freien Universitaet |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.