
Bellwoodilabrus landinii, Bannikov & Carnevale, 2010
|
publication ID |
https://doi.org/ 10.5252/g2010n2a2 |
|
persistent identifier |
https://treatment.plazi.org/id/038E87F8-A961-B17A-98A2-FD10FB4AFFC0 |
|
treatment provided by |
Felipe |
|
scientific name |
Bellwoodilabrus landinii |
| status |
n. gen., n. sp. |
Bellwoodilabrus landinii n. gen., n. sp. ( Figs 1-4)
HOLOTYPE. — MCSNV IGVR 71214 /71215, part and counterpart, complete articulated skeleton, 43.5 mm total length (TL), 34 mm standard length (SL). Only known specimen.
ETYMOLOGY. — It is our pleasure to name this species in honour to the Italian paleontologist Prof. Walter Landini, a mentor to one of us (GC), valuable colleague to all of us, in recognition of his contribution to the study of fossil labrids and other Tertiary fishes.
TYPE LOCALITY AND HORIZON. — Monte Bolca locality, Pesciara cave site; Early Eocene, late Ypresian, middle Cuisian, SBZ 11, Alveolina dainelli Zone (see Papazzoni & Trevisani 2006).
DIAGNOSIS. — As for the genus.
MEASUREMENTS ( AS PERCENTAGE OF SL). — Head length (HL) = 31; maximum body depth = 41; caudal peduncle depth = 17; snout length = 10; orbit diameter = 7.5; lower jaw length = 16; distance between tip of the snout and first dorsal-fin spine = 42; distance between tip of the snout and first dorsal-fin ray = 75; distance between tip of the snout and anal fin = 66; distance between pelvic and anal fins = 26; dorsal-fin base = 53; spinous dorsal-fin base = 33; anal-fin base = 30; length of the first dorsalfin spine = 8; length of the longest dorsal-fin spine = 13; length of the first anal-fin spine = 9; length of the third anal-fin spine = 16; length of the pelvic-fin spine = 12; length of the longest caudal-fin ray ≈ 28.
DESCRIPTION
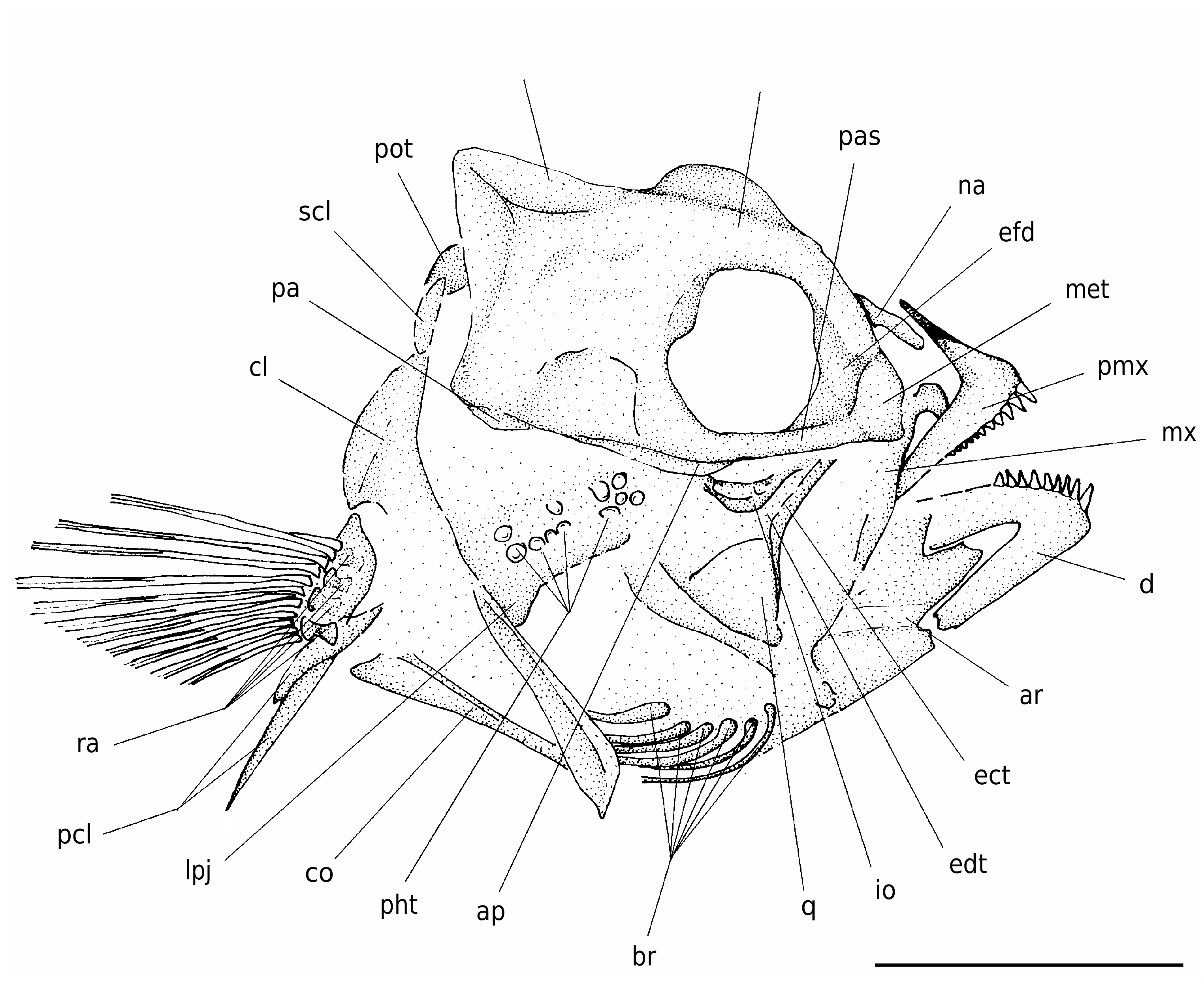
The body is moderately elongate, nearly ovoid in outline and laterally compressed, with a short and deep caudal peduncle ( Figs 1; 2 View FIG ). The head is rather large, contained more than three times in SL. The head is relatively deep, its maximum depth almost equals its length; the snout is moderately elongate, contained three times in HL. The mouth is terminal, nearly horizontal, with a moderately large gape. The posterior corner of the lower jaw is situated below the anterior third of the orbit.
The neurocranium is relatively deep and characterized by a developed bony relief ( Fig. 3 View FIG ). The vomer is massive and compressed anteroposteriorly. The mesethmoid has a nearly vertical anterior profile. The ethmoid-frontal depression (= frontal recess; see Gomon 1997) is moderately developed. The frontals are characterized by a prominent median bony flange projecting above the orbit. There is a well-developed supraoccipital crest that extends anteriorly to the frontal region. The posterior margin of this crest has a nearly vertical profile. A ridge formed by the epioccipital, parietal and frontal, which terminates along the posterior sector of the frontal region, appears to be present. The parasphenoid is robust, nearly linear, and bears two processes, the adductor process to which the adductor arcuus palatini muscle attaches and a posterior pharyngeal apophysis to which in origin attached an articular process of the third pharyngobranchial; the adductor process (see Rognes 1973; Rosen & Patterson 1990) is quite elongate, while the pharyngeal apophysis has a rounded ventral profile.
The nasal is a simple bar-like moderately elongate bone that is closely associated to the frontal; this bone is slightly curved and is characterized by a tube for the supraorbital cephalic laterosensory canal oriented through its long axis. What appear to be the third and part of the fourth infraorbital bones can be recognized. These bones are platelike, with a slightly expanded planar bony flange ventrally.
The premaxilla has a well-developed ascending process; as in other labrids, the articular process of the premaxilla appears to be coalescent with the ascending process ( Rognes 1973; Rosen & Patterson 1990). The development of the ascending process of the premaxilla is possibly correlated with the development of the median ethmoid- frontal depression, which partially encloses the distal ends of the ascending processes of the premaxillae, thereby forming the ascending process fossa (see Rognes 1973; Russell 1988; Rosen & Patterson 1990). The horizontal alveolar process is longer than the ascending process and forms an angle of about 80° with it. The posterior end of the alveolar process is slightly curved ventrally. The premaxilla bears a single row of progressively smaller conical teeth, which are slightly blunt apically. The maxilla is only partially preserved, represented by the dorsal process (see Van Hasselt 1979), and by part of the anteroventrally curved main axis. The lower jaw is moderately deep, its length slightly exceeds the half of HL. The dentary has a nearly triangular configuration. The symphysis of the dentary is oblique and relatively short. The oral border of the dentary possesses strong and apically blunt conical teeth arranged in a single row. The morphology of the angular and articular is unclear.
The bones of the suspensorium are only partially preserved. The main shaft of the hyomandibula is obliquely oriented. The quadrate is subtriangular in outline. Both the ectopterygoid and endopterygoid are narrow and elongate. The palatine is not preserved.
The opercular bones are not clearly recognizable. The preopercle seems to be slightly curved; there is no evidence of serrations along the posterior margin of this bone.
There are six sabre-like branchiostegal rays, the first of which is narrower and weaker. The large part of the branchial skeleton is badly preserved and the elements are difficult to recognize. The pharyngeal jaws are certainly hypertrophied; a diarthrosis between the upper pharyngeal jaws (third pharyngobranchials) and the pharyngeal apophysis of the parasphenoid was probably present, and the lower pharyngeal jaws (fifth ceratobranchials) are definitely fused. Although the lower pharyngeal jaw probably had some form of physical contact with the cleithrum, there is no evidence of a cleithral articular process fossa, implying that a true synovial joint ( Liem & Greenwood 1981) was probably absent in origin. The pharyngeal teeth are relatively large; those of the lower jaw are rounded
B
and molariform, whereas upper pharyngeal teeth mostly have roundly conical crowns.
The vertebral column consists of 24 (9+15) vertebrae, including the urostyle. The axis of the vertebral column is slightly curved ( Fig. 2 View FIG ). The vertebral centra are rectangular to subrectangu- lar, longer than high. The neural spine of the first vertebra is expanded anteroventrally and is slightly shorter than those of the other vertebrae, but not strongly reduced as in many labrid taxa (see Russell 1988; Gomon 1997). Well-developed parapophyses are clearly recognizable in the three posterior abdominal vertebrae. Prominent neural prezygapophyses characterize the posterior abdominal vertebrae. The haemal spines of the anterior caudal vertebrae are slightly stronger and longer than the opposite neural spines. There are seven pairs of slender pleural ribs. Thin epineurals articulate on the parapophyses of the seventh and eighth abdominal vertebrae.
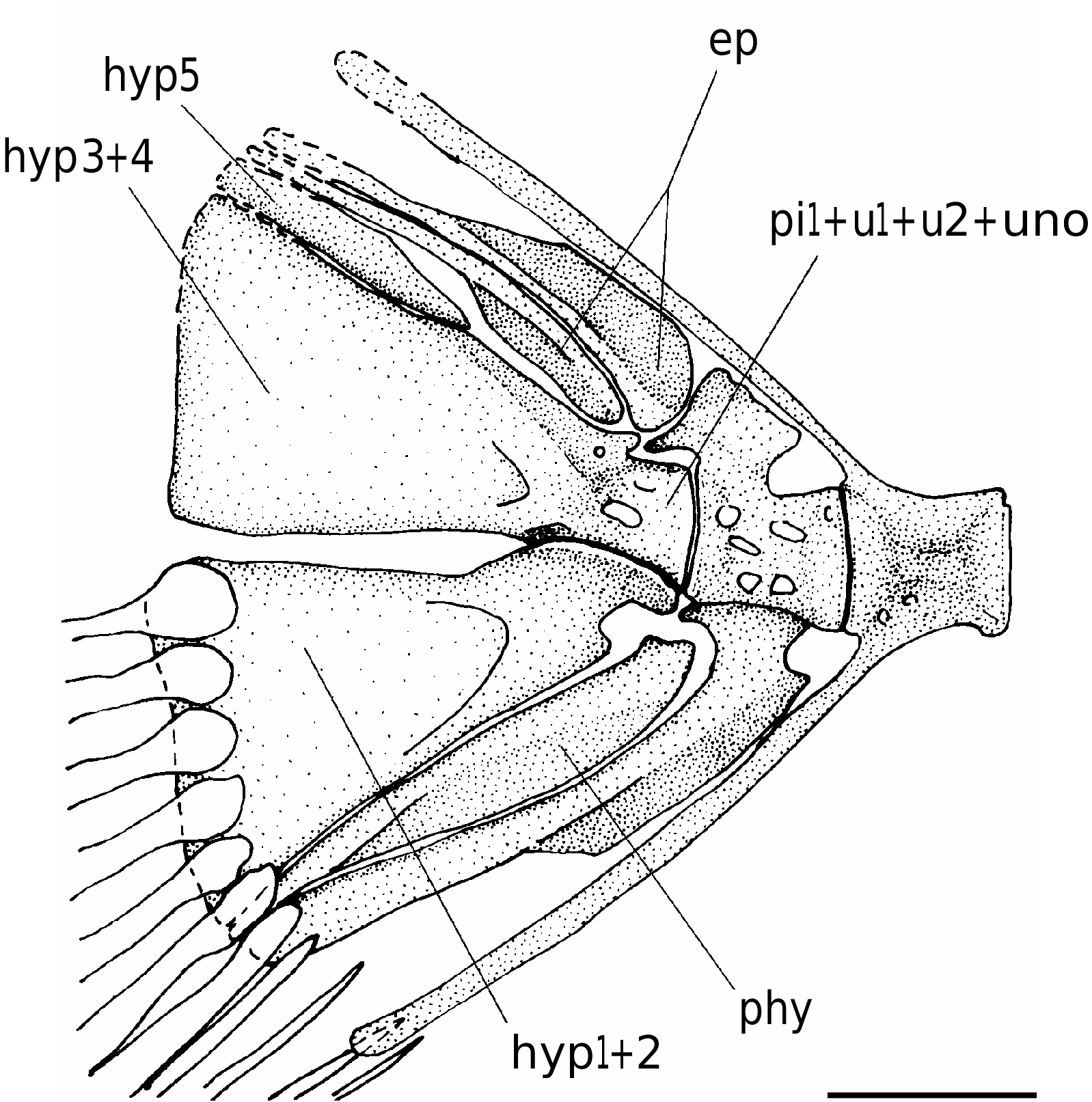
The caudal skeleton has the typical labrid structure ( Fig. 4 View FIG ). The urostyle is characterized by the fusion of the first preural centrum to the first and second ural centra; it is completely fused to the uroneural, forming a solid block ankylosed to the fused hypurals 3 and 4. The hypurals 1 and 2 are fused into a roughly triangular hypaxial hypural plate. The reduced fifth hypural (“hypurale minimum”; Monod 1968) is autogenous. There is a deep hypural diastema between the epaxial and hypaxial plates. The parhypural is autogenous; it does not reach the terminal half-centrum and apparently lacks the parhypurapophysis. The haemal spine of the penultimate vertebra is autogenous, while that of the antepenultimate vertebra is fused to the centrum. The neural spine of the penultimate vertebra consists of a short rhomboid crest. There are two epurals, the first of which is longest and more expanded anteroposteriorly. The caudal fin is moderately large and rounded; it consists of 14 principal rays (I,6-6,I), plus five upper and four lower procurrent rays.
There is a single thin supraneural, which is nearly straight ( Fig. 2 View FIG ). The supraneural is obliquely oriented, extending posteriorly in front of the distal tip of the second neural spine. The dorsal fin is rather elongate, its base extension exceeds half of the body length. The dorsal fin is continuous and originates just above the fifth vertebra. The bases of the two anterior closely associated pterygiophores are located between the neural spines of the second and third vertebrae. The dorsal fin contains nine slender and pungent spines and 11 soft rays, supported by 19 pterygiophores. The spines gradually increase in length from the first to the third, and succeeding spines are almost equally long. The first dorsalfin spine is in supernumerary association on the first dorsal-fin pterygiophore. The pterygiophores of the spiny portion of the dorsal fin are characterized by a narrow and long main shaft that is bordered posteriorly by a medially flattened bony blade; each of these pterygiophores bears a spine in supernumerary association, and the eleventh pterygiophore additionally bears serially associated first soft ray.
The anal fin originates just below the third caudal vertebra and ends at the level of the penultimate vertebra. The anal fin consists of three anal spines and nine branched rays, supported by 10 pterygiophores; the first two anal-fin spines are supernumerary. The spines are strong and show a progressive increase in size. The longest analfin rays are slightly longer than the third anal-fin spine. The first anal-fin pterygiophore is relatively long and sturdy; its shaft is inclined at an angle of about 73° to the body axis and inserts behind the first haemal spine.
The pectoral girdle is rather robust ( Fig. 3 View FIG ). The posttemporal and supracleithrum are poorly preserved. The cleithrum is large and elongate. The ventral postcleithrum is slender and rib-like, whereas the dorsal one is short and wide. The coracoid is relatively narrow, characterized by a long and tapered postcoracoid process. The scapula is difficult to recognize. The radials are incompletely preserved, but all the elements appear to be orientated in the same direction. The base of the pectoral fin is moderately wide and obliquely oriented, located just under the fourth and fifth vertebrae, close to the midpoint between the vertebral column and the ventral profile of the body. The pectoral fin contains 12 relatively short rays. The first ray is very short and not branched.
The basipterygium is a long and narrow bone ( Fig. 2 View FIG ). The pelvic fin contains one spine and five rays. The fin inserts at the same level as the pectoral fin insertion. The pelvic-fin rays are longer than the pectoral-fin rays.
Thin cycloid scales cover the entire body, including the bases of the anal and spinous dorsal fin. Some scales reveal few indistinct radii in the basal field. The lateral line has a smoothly curved contour, nearly parallel to the dorsal body profile; it is interrupted under the third soft dorsal-fin ray ( Fig. 2 View FIG ).
A new labrid fish ( Teleostei, Perciformes ) from the Eocene of Monte Bolca

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.