
Listriodon splendens von Meyer, 1846
|
publication ID |
https://doi.org/ 10.5281/zenodo.5377612 |
|
persistent identifier |
https://treatment.plazi.org/id/038987C1-F346-FFFE-9AFF-FBE4FB2ACDFA |
|
treatment provided by |
Marcus |
|
scientific name |
Listriodon splendens von Meyer, 1846 |
| status |
|
Listriodon splendens von Meyer, 1846
The middle Miocene suid, Listriodon splendens , has been known for well over a century, and fossils assigned to this species have been recorded from over a hundred localities in Europe, Asia and Africa yet complete skulls are extremely rare. Measurements of the dentition of French Listriodon splendens are given in Table 3.
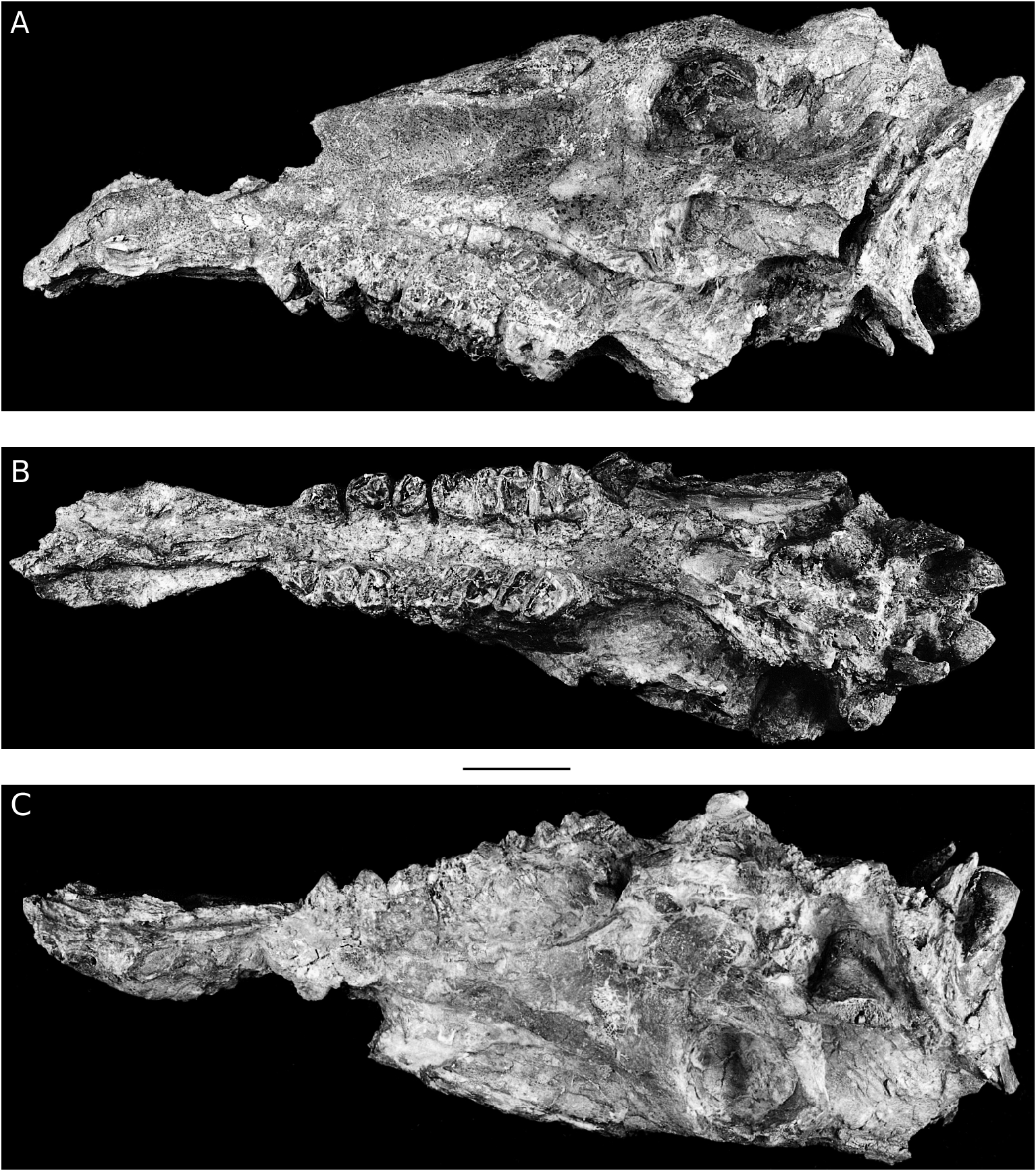
CRANIUM OF LISTRIODON SPLENDENS FROM TORIL, SPAIN
The Toril cranium (TO 3B, 129) ( Figs 12 View FIG ; 13 View FIG ), presently stored at the Museo Nacional de Ciencias Naturales, Madrid, but belonging to the Museo de Paleontologia , Zaragoza, consists of a virtually complete cranium which has been crushed laterally and slightly obliquely. It lacks the anterior dentition back to and including the
P1 but the cheek tooth series (P2-M3) on both sides are present, though the teeth are heavily worn and damaged by cracking and expansion. The premaxillary margins of the external nares are preserved, but the anterior parts of the nasal bones have broken away. Both zygomatic arches have been damaged and much of the squamous part of the occipital bone is missing so that the nuchal crest is not preserved. Interpretation of the basicranium is rendered difficult because of the crushing that it has undergone, but enough is preserved for the major structures to be observed. Cranial sutures are not visible in this specimen which was evidently almost senile when it died, judging from the stage of wear of the dentition.
Palatal view
In palatal view, the anterior part of the snout is spatulate, being narrowest behind the canines
Length i1 i2 i3 p2 p3 p 4 m 1 m 2 m 3 i1 i2 i3 p2 p3 p 4 m 1 m 2 m 3 Breadth and widest between the canine and I3. There is a substantial diastema (c. 5 cm) between the posterior margin of the canine alveolus and the anteri- or edge of the P2 and a shorter diastema (12 mm) between the anterior edge of the canine alveolus and the posterior margin of the alveolus for I3. The edge of the palate between the canine and the P2 is delimited by a narrow crest of bone and there is no sign of alveoli for P1.
The posterior nares are located far back (33 mm) behind the M3. The pterygoid process of the palatine is well developed, as in Sus Linnaeus, 1758 . The vertical portions of the pterygoid and the pterygoid process of the sphenoid are poorly preserved, but appear to have been similar to those of Sus .
In Sus there is a substantial space between the pterygoids and the tympanic bullae, but in the Toril cranium, the anterior portion of the bullae terminate close to the pterygoids. This is due partly to a relative increase in the size of the bullae, and partly due to their inclination at a marked angle. Whereas in Sus the bullae are oriented almost vertically, such that in lateral view the angle between their distal edge and the paramastoid process is narrow (c. 15°), in Listriodon the angle is much wider (100°), the bullae pointing towards the palate. In addition, the bullae in Sus are ovoid with a low but sharp anterior crest, whereas in Listriodon the anterior part of the bullae including the crest are much expanded. The morphology and inclination of the tympanic bullae in Listriodon is close to that described for Lopholistriodon kidogosana Pickford & Wilkinson, 1975 by Pickford (1986).
The paramastoid process in Sus is long and oriented vertically, bending slightly backwards towards their tips, and they thus extend well below the occipital condyles, terminating in the same plane as the occlusal surface of the cheek teeth. In Listriodon in contrast, the processes are shorter and terminate just below the occipital condyles and well above the occlusal plane of the cheek dentition. They also point more distally. In these features, Listriodon splendens resembles Lopholistriodon kidogosana .
The temporal condyles are located high above the occlusal plane of the cheek teeth, much as in Sus . However, the ventral temporal crest is much closer to the temporal condyle, especially its median end, and in this Listriodon and Lopholistriodon are similar to each other and both differ from Sus .
Lateral view
The anterior nares of the Toril Listriodon extend as far back as the anterior margin of the canine, whereas in Sus they terminate further forwards, over the I3. The Retama snout of Listriodon retamaensis n. sp. has shorter external narial margins, more like the situation in Sus .
In the Toril cranium the infraorbital foramen is located above the P3 whereas in Sus it is usually located further back over the M1. The anterior end of the crest on the maxilla which separates the fossa for the levator rostri muscles above from the depressor rostri muscles below is located above the P4. The zygomatic arches sweep backwards at a gentle angle with no sign of a pre-zygomatic shelf. The preorbital fossa, which in Sus is well developed distally, forming an overhanging ledge in the lachrymal bone and a depression in the malar bone, is poorly developed in Listriodon . In the Toril lachrymal, there is a shallow depression with no overhang, and in the malar there is no trace of a fossa. In Sus the rhinarial musculature associated with rooting behaviour originates in the preorbital fossa, from which we infer that in Listriodon , the rhinarial musculature was probably relatively weakly developed. The rounded profile of the anterior part of the snout in Listriodon also indicates that there was not a major muscle mass leading forwards towards the rhinarium. In Sus , in contrast, the snout is almost square in profile, with the side of the face possessing a wide groove along which pass the muscles and tendons of the rhinarial musculature which insert into the rostral bone. In Sus , above the third molar there is a triangular process projecting ventrally from the base of the prezygomatic crest. This process separates the lower part of the facial aspect of the maxilla (where the depressor rostri musculature originates) from its maxillary recess. In Listriodon there does not appear to be an analogous process, there being no clear separation between the maxillary recess and the ventral surface of the root of the zygomatic arch.
In Listriodon the zygomatic arch is relatively much deeper than it is in Sus . Taking the greatest diameter of the orbit as a reference, the depth of the zygomatic arch taken lateral to the maxillary recess is about 61% of this diameter in Sus (26 mm / 43 mm), whereas in Listriodon the depth of the zygomatic measured in the homologous position is over 100% the orbital diameter (67 mm / 53 mm).
There appears to be no lachrymal foramen in Listriodon , in contrast to the double foramen located on the margin of the orbit in Sus . Lopholistriodon likewise has no lachrymal foramina, but Libycochoerus Arambourg, 1961 and Kubanochoerus Gabunia, 1955 , each possess a single foramen. The outline of the orbits in the Toril specimen is more squared off than are the orbits of most suids. In particular the corner of the orbit between the anterior and ventral margins forms almost a right angle on both sides of the skull, but it is not clear whether this angulation has been enhanced by or is due solely to plastic deformation and crushing.
The malar of Sus possesses no foramina, but both malars in the Toril Listriodon cranium possess a prominent foramen below and anterior to its triangular temporal process. As far as we are aware, such a foramen has not previously been reported to occur in Suidae , and its presence in Listriodon may well be a synapomorphy of the subfamily. In Listriodon from Toril, the temporal process of the malar forms a prominent spine reaching upwards and backwards towards the supraorbital process, thereby reducing the extent of the post-orbital opening (c. 13 mm), which, in Sus , is wide open (c. 25 mm in a much smaller skull used for comparison). The inferior margin of the zygomatic arch of the Toril cranium has been broken, revealing that it is extensively invaded by sinuses. The orbits of the Toril cranium lie entirely behind the third molars, their position being close to the condition observed in Sus scrofa Linnaeus, 1758 .
In Listriodon the external accoustic meatus is high, as in Sus , opening about half the distance between the base of the occipital condyles and the nuchal crest. The detailed architecture of the bones surrounding the temporal fossa is difficult to make out on account of the crushing and plastic deformation that the Toril specimen has undergone. However, in overall shape, it appears
Upper dentition Length Breadth P3 16.9 15.6 P3 15.5 11.5 P4 14.3 16.6 M1 17.4 17.5 M2 19.8 19.7 M3 25.4 20.9 Lower dentition Length Breadth p2 14.8 8.4 p3 17.0 9.6 p3 15.9 10.1 p4 16.4 13.3 p4 16.7 12.9 m 1 16.4 12.5 m 1 17.1 13.1 m 2 20.8 16.3 m 3 31.9 19.6
that this part of the cranium was not markedly different from that of Sus . The parietal and temporal crests are well developed, but the nuchal crest is missing in the fossil. The frontal contains extensive sinuses as shown by a break above the right orbit.
From the above description it is evident that the Toril cranium is close in general morphology and proportions to that of Sus scrofa . Yet there are several important differences between the skulls of the two genera. The preorbital fossa in the lachrymal and malar, which in Sus is highly developed, is reduced or absent in Listriodon . This suggests that the rooting habit was not important in Listriodon . There is no lachrymal foramen outside the orbit in Listriodon , whereas in Sus there is usually a double foramen outside the orbit. Unique to Listriodon is the presence of a large foramen near the temporal process of the malar. This process is well developed and forms a prominent spine which reaches upwards towards the supraorbital process thereby reducing the gap between these two bones. In Sus scrofa , the temporal process of the malar barely extends above the zygomatic process of the temporal bone, whereas in Listriodon it projects well above it.
Pickford M. & Morales J.
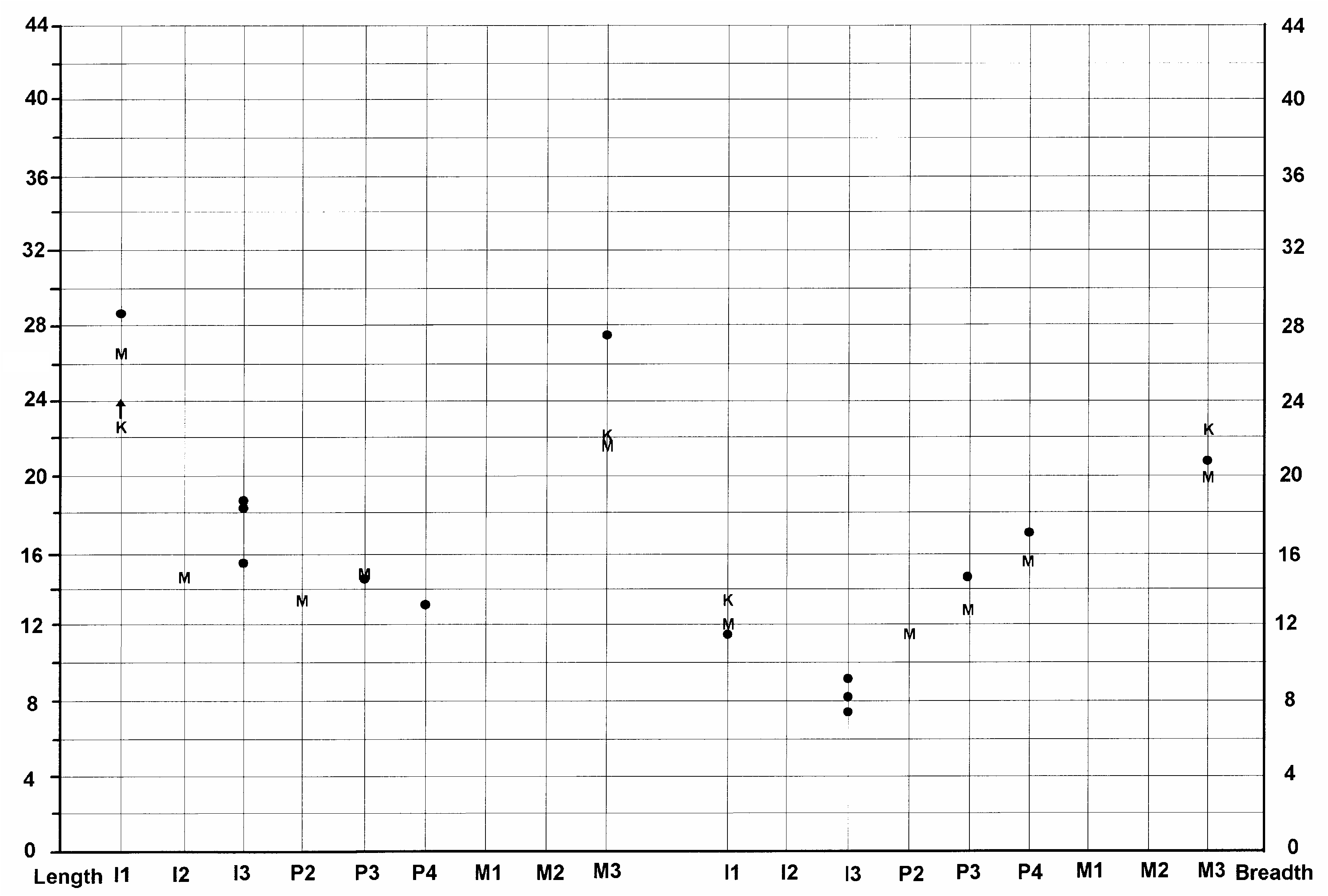
FIG. 14. — Size variation (in mm) of the upper dentition of Listriodon splendens von Meyer, 1846 . Symbols and abbreviation:, maximum and minimum measures; l, range of variation; T, Toril specimens.
In the neurocranium, the most obvious differences between Sus and Listriodon are the enlarged tympanic bullae in the latter genus which point more anteriorly than in Sus and the short paramastoid processes which terminate well above the occlusal plane of the cheek dentition. In Sus the paramastoid processes are long and terminate on a level with the occlusal surface of the cheek teeth. In view of the fact that some of the movements of the mandible are controlled by muscles which originate at the ends of the paramastoid processes, the major differences between these two suids indicate important functional differences in mastication.
In palatal view the most obvious differences between the Toril Listriodon and Sus , apart from the dentition, are the spatulate anterior part of the snout, the elongated diastemata either side of the canine and the retired posterior nares which open up well behind the third molars.
Upper dentition ( Table 4)
In the Toril cranium the three upper incisor alveoli form a closed series separated by a short diastema from the canine alveolus. The canine alveolus is small and opens downwards, forwards and laterally, and is bordered on its dorsal side by a low canine jugum. The size of the alveolus, its orientation and the low, non-projecting jugum indicate that this individual was probably a female. Male listriodonts usually possess large projecting canine juga forming canine flanges and much larger canines that emerge from the end of the canine flange and then flare sideways before curving laterally and upwards. In the Toril cranium there is a long diastema behind the upper canine and there is no sign of alveoli for the first upper premolar which appears either to have been a rudimentary tooth or to have been suppressed altogether. By way of contrast, the Retama female snout of Listriodon retamaensis
Length i1 i2 i3 p2 p3 p 4 m 1 m 2 m 3 i1 i2 i3 p2 p3 p 4 m 1 m 2 m 3 Breadth
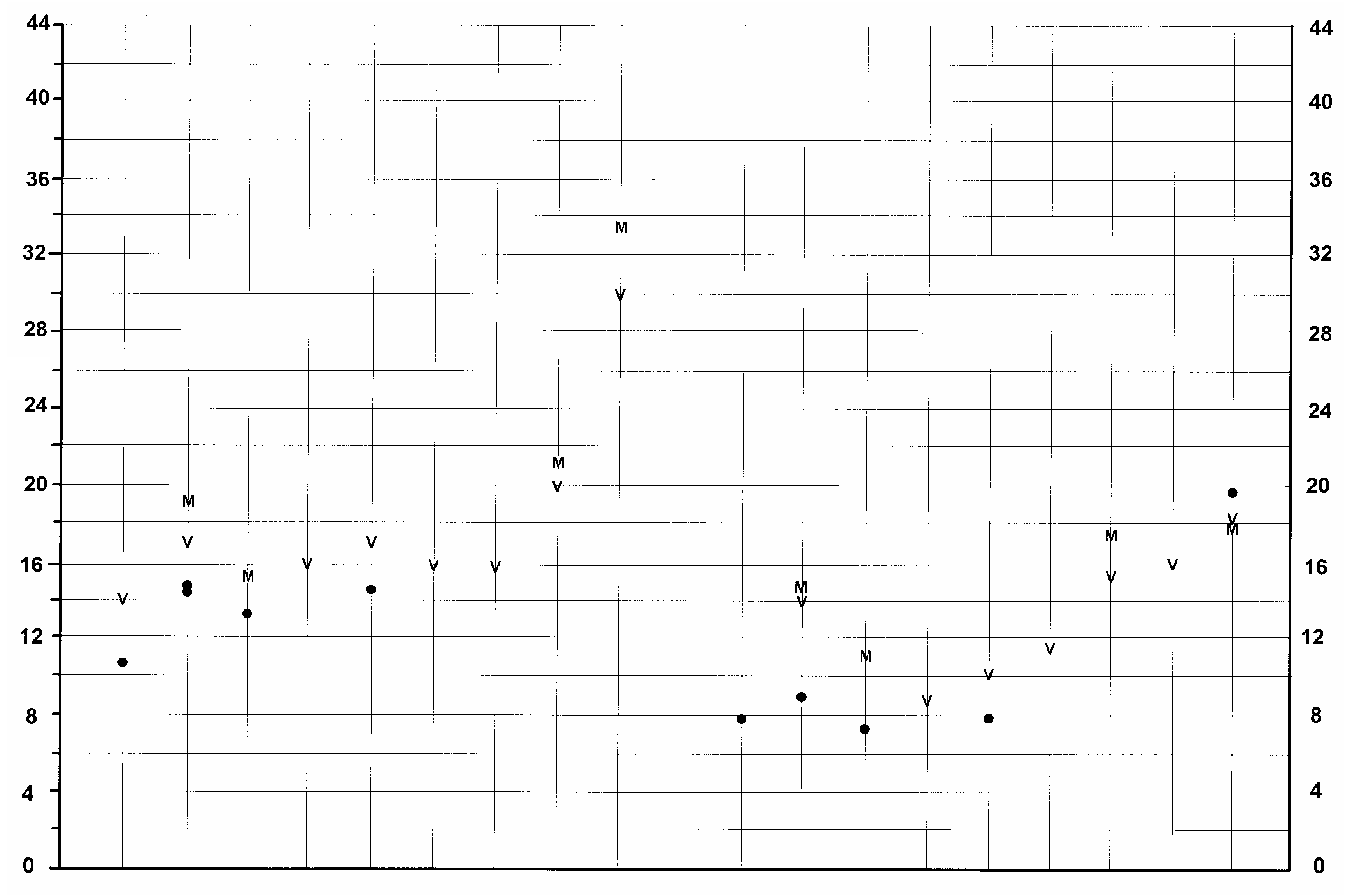
FIG. 15. — Size variation (in mm) of the lower dentition of Listriodon splendens von Meyer, 1846 . Symbols and abbreviation:, maximum and minimum measures; l, range of variation; T, Toril specimens.
n. sp. has a double rooted P1 situated midway between the canine and the P2. In the Toril cranium the P2-M3 form a closed dental series.
The cheek teeth in the Toril skull are heavily worn and have suffered damage which means that accurate dimensions are difficult to obtain. The measurements given in the table overestimate most dimensions, because the teeth have cracked and “expanded”. In lateral view, the occlusal surface of the cheek teeth curves gently ventrally as one progresses from the molars towards the premolars.
OTHER LISTRIODON SPLENDENS FOSSILS FROM TORIL ( TABLES 4; 5)
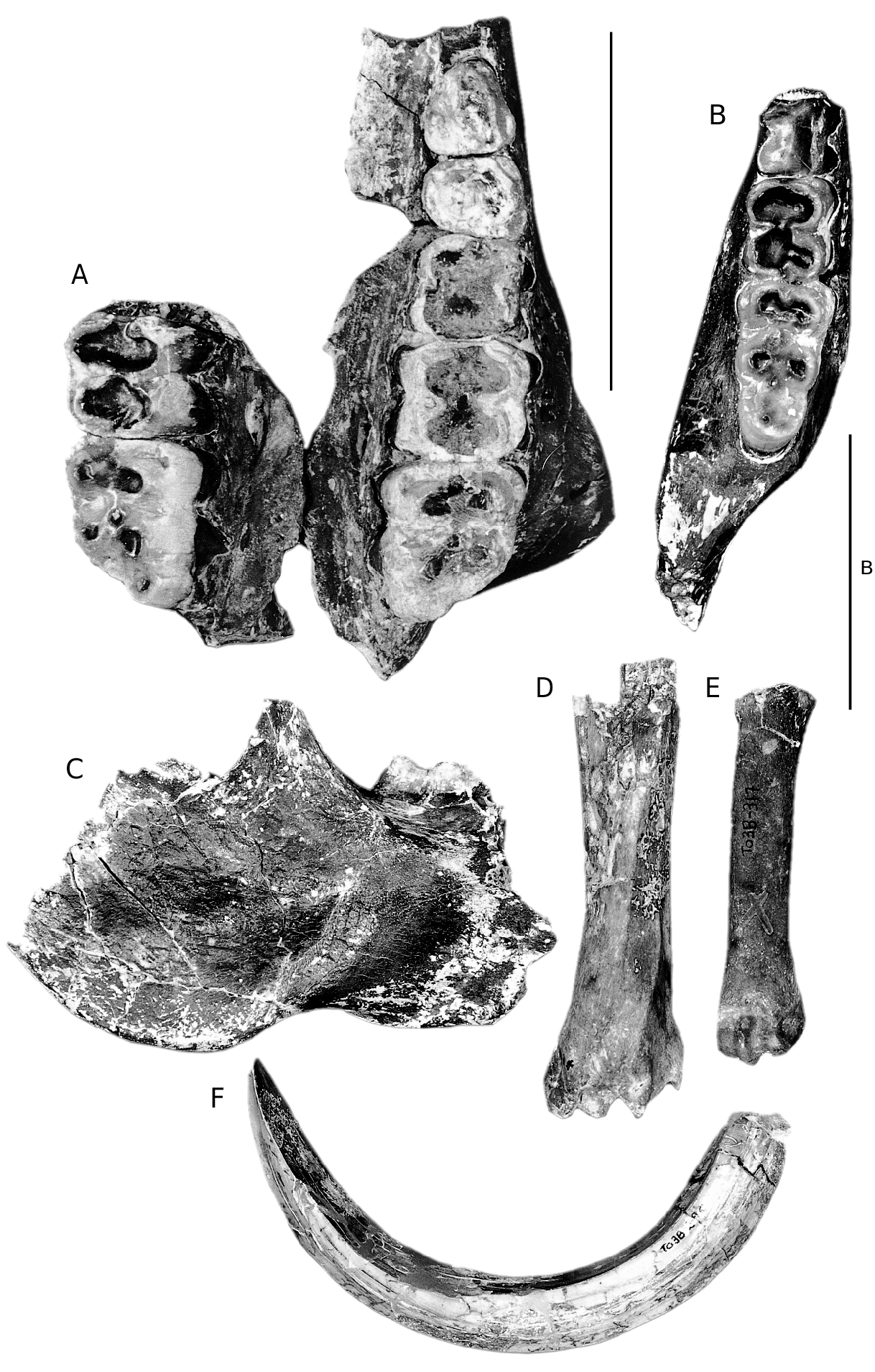
In the collection from Toril, there are several isolated listriodont teeth and a few post-cranial bones ( Figs 1 View FIG D-G; 2D-F). There are three upper central incisors in the sample, but they are either broken or extremely worn. TO 3B, 337 and 334
are right I1s, both of which are so worn that only a little enamel remains on the labial surface and tiny patches lingually. Both individuals suffered from heavy abrasion of the root on the labial side indicating that the gingiva had retracted and exposed the cervical part of the root to abrasion from vegetation and also the lips. In TO 3B, 337 the abrasion surface extends for c. 5 mm from the crown-root margin, while in specimen 334, the surface extends c. 10 mm. In both specimens the abraded surface of the roots is bordered by a series of shallow, rounded, sharp-edged depressions up to 1 mm in diameter, probably caused by a periodontal disease. TO 3B, 13 is the unworn germ of a left I1 with an incompletely formed root which has been crushed buccolingually. It lacks the mesial corner of the crown which has been replaced with plaster of Paris. The occlusal edge of the crown is marked by many small indentations producing a beaded
Pickford M. & Morales J. FIG. 16. — Size variation (in mm) of the upper dentition of Eurolistriodon adelli Pickford & Moya Sola, 1995 . Symbols:, maximum and minimum measures; l, range of variation.
effect. The distal margin of the crown overhangs the root, and there is a prominent lingual cingulum. The central lingual pillar is poorly developed.
TOR 3A 223’98 is an unworn right P3. The occlusal outline of the tooth is triangular with rounded corners. The apex of the main cusp lies over the centre of the tooth and it has a p r o m i n e n t c r e s t d e s c e n d i n g l a b i o d i s t a l l y towards the cervix. A second shorter crest runs mesiolingually. The anterior surface of the main cusp has a wear facet oriented transversely, caused by its lower antagonist, as described by Leinders (1977). There is a low distolingual cusp separated from the main cusp by a valley which opens distally into a low shelf bordered by the distal cingulum. There is also a low cingulum anteriorly. There are three roots, the dis- t a l p a i r b e i n g f u s e d l a b i o l i n g u a l l y i n t o a wall-like structure.
TOR 3A 508’98 is a lightly worn left dM4 with a slightly trapezoidal, almost rectangular occlusal outline. The four main cusps are arranged into two lophs with a continuous crest joining the main cusps to each other located slightly anterior to them. Each of the main cusps has a subsidiary crest running from its tip transversely towards the midline of the tooth. The paracone has a “zygodont” crest running distally from its tip into the median transverse valley. There is a prominent anterior, labial and distal cingulum.
TOR 3B 438 is a right i1. It has an almost bilaterally symmetrical crown with a prominent lingual pillar and low lingual cingular ridges. The root is straight and robust. TO 3B, 201 is a right i 2 in light wear. It has a scooped out distal margin, a well developed central lingual pillar with a bulbous base and the lingual mesial margin is raised into a crest. TOR III (33.234) is a left i3 with a prominent wear facet apically. Its root
Length i1 i2 i3 p2 p3 p 4 m 1 m 2 m 3 i1 i2 i3 p2 p3 p 4 m 1 m 2 m 3 Breadth
FIG. 17. — Size variation (in mm) of the lower dentition of Eurolistriodon adelli Pickford & Moya Sola, 1995 . Symbols:, maximum and minimum measures; l, range of variation.
narrows rapidly from cervix towards its apex which is curved. The crown is triangular in lingual view and sits obliquely on its root. Mesially it is missing a small part of the crown. It possesses a lingual cingulum and there is a weakly developed lingual pillar. There is also a cingulum on the distal part of the labial surface of the crown. TOR 3B 322 is also a left i3 but it is appreciably small- er than the previous specimen. In RET 338, i3 possesses a triangular crown posed obliquely on its root. The lingual cingulum and pillar are weakly developed and there is no labial cingulum. The distal scoop is well expressed and has a small wear facet cutting its distal margin. The presence of two such different i3s from the same site poses a problem. Either these teeth were extremely variable, or they may be sexually dimorphic.
TOR 3B 290 is a small section of a lower canine of a male individual. TO 3B, 295 is a complete lower canine of a male individual ( Fig. 2F View FIG ). It has enamel on the lingual and labial surfaces but there is none on the distal surface. The distal surface has a broad central ridge, but near the apex of the tusch, this ridge has been abraded away by wear against the upper canine. In section the canine is scrofic, with a slightly larger lingual (23.2 mm) than labial surface (20.8 mm). TOR 3B 197 is a lightly worn but broken right lower molar and TOR III (33.233) is the unerupted crown of a left M2.
TOR 3A 1335’98 is a broken right p4, lacking the buccal and anterior half of the main cusp. The innenhugel is robust and well offset from the main cusp, being joined to it by a transverse crest. Distally there is a low but robustly constructed posterior accessory cusp accompanied by a distal cingulum which fades out labially and lingually as it approaches the main cusp.
The cheek teeth in the Toril cranium are very large, plotting at the upper end of the range of variation of Listriodon teeth, or even falling beyond the known range of variation in the case of the P2 (Figs 14; 15). However, in view of the cracking and expansion that the teeth in the skull have undergone, we should perhaps reduce the measurements by 1 or 2 mm in order to obtain a more accurate estimate of the original dimensions of the teeth. Even if we do this, the material still plots out at the large end of the size range of listriodont teeth (Figs 14; 15). The P3 and P2 have incipient metacones, which, when combined with the large dimensions of the cheek teeth suggests that the Toril species should be identified as Listriodon splendens major Roman, 1907 . The greatly expanded mesiodistal dimension of one of the i3s confirms this identification.
Postcranial bones ( Table 5)
There are several suid postcranial elements from Toril. The left talus TOR 3B 107 is large, plotting in the upper part of the range of distribution of Listriodon tali. The measurements of other bones are given in Table 5.
DISCUSSION
There has been a long-running debate ( Van der Made 1997b) about the validity of the genus Bunolistriodon , which the Toril cranium helps to resolve. There have been two main points of view in the literature. The first is that Bunolistriodon is so close to Listriodon that the two should not be separated at the generic level ( Gabunia 1973; Leinders 1975; Pickford 1986). The other is that the dentition and skulls are widely divergent and that this warrants their generic separation. However, the erroneous view that the skulls of Bunolistriodon and Listriodon are markedly different is based on a combination of two factors. Firstly there has been a lack of comparative material, the skulls of both supposed genera being rare and often crushed or distorted, and secondly, many of the skull characters claimed to distinguish Bunolistriodon from Listriodon do not occur in the former genus, but were drawn from a suid skull from Libya ( Libycochoerus massai Arambourg, 1961 ) that belongs to a different subfamily, the Kubanochoerinae . The importance of the Toril Listriodon cranium is that it reveals that there are no major differences in cranial architecture between the European skulls hitherto assigned to Bunolistriodon and those assigned to Listriodon . The differences observed are of lesser degree than those that separate two species of the genus Sus ( S. scrofa and S. verrucosus Müller, 1840). On this basis one should seriously question whether the continued use of these two genera within Listriodontinae is warranted. The European dentitions assigned variously to Bunolistriodon and Listriodon form a morphological continuum from buno-lophodont forms such as occur in the species L. retamaensis n. sp. and L. latidens on the one hand and more lophodont forms such as occur in L. splendens on the other. There is no clear separation into two morphometric groups. From this it is concluded that Bunolistriodon is not generically separable from Listriodon and is therefore a junior synonym of it. In general features the skull of Eurolistriodon adelli ( Pickford & Moya Sola 1995) , even though the only known specimen is badly crushed, is typically listriodont, and were it not for the precocious loss of P1 and P2, the exaggerated development of canine flanges in males which extend like a horn as a separate projection beyond the canine alveolus, together with its bunodont (almost crystodont) dentition, it would also fit within the genus Listriodon . The form of the canine flange is so different from that of other Listriodon species , however – the canines emerge from the ventral side of the canine flange, not at its end – that when added to the dental and postcranial differences enumerated above, it means that at Els Casots we are in the presence of a genus distinct from Listriodon . The cranial differences are reflected in the postcranial skeleton, with Eurolistriodon possessing elongated, relatively gracile metapodials and Listriodon shorter, more robust ones.
A second area of divergent opinions in suid systematics is whether the kubanochoeres (subfamily Kubanochoerinae ) should be subsumed into
TOR 3B 107, talus External length (Lext) 47.3 Internal length (Lint) 43.3 Proximal transverse diameter (DTp) 24.0 Length in the middle (Lm) 38.3 Distal transverse diameter (DTd) 28.8 Width of cuboid facet (d) 12.0 Diameter of tibial pulley (R) 28.3 TO III (33.236), proximal end of a third metacarpal Proximal transverse diameter (Dtp) 34.0 Proximal antero-posterior diameter (DAPp) 29.3 TO 3B, 333, distal tibia Mesiolateral diameter (Dtd) 35.0 Anteroposterior diameter (DAPd) 32.0 TO 3B, 317, left fourth metatarsal Length (L) 104.2 Distal mesiolateral diameter (Dtd) 21.0 Distal anteroposterior diameter (DAPd) 21.8 TO 3B, 30, distal end of a second or third metapodial Mesiolateral diameter (Dtd) 23.6 Anteroposterior diameter (DAPd) 23.2 TOR 3B 250, juvenile second phalanx lacking the proximal epiphysis Distal breadth (Dtd) 13.5 Distal height (DAPd) 10.2 TOR 3B 108, terminal phalanx Length (L) 34.2 Anterior height (DAP) 23.0 Anterior breadth (Dtp) 13.5
Listriodontinae as proposed by Van der Made (1996, 1997b) or whether they should not, a position maintained by Pickford (1986, 1993). Van der Made, on the basis of a few characters such as the enlarged I1, considers that these two subfamilies should be united together. However, Pickford (1986, 1993) has pointed out numerous fundamental differences between the skulls and dentitions assigned to these two groups. The Toril cranium, by providing further information that was hitherto not available, serves to underline the fundamental differences between these two suid lineages.
Listriodonts do not possess lachrymal fossae on the orbital margin, whereas the skulls of Libycochoerus massai and Kubanochoerus gigas (Pearson, 1928) do. The loss of these foramina appears to be a synapomorphy of the Listriodontinae , as they are unknown in any of the available Listriodon skulls including those from China and the skull of Lopholistriodon kidogosana from Kenya, but are present in all other known lineages of suids. Listriodon possesses a prominent foramen on the malar bone, just below the triangular apex of its zygomatic process. Such a foramen is unknown in other suids, and it probably represents a synapomorphy of the subfamily. The tympanic bullae of Listriodon and Lopholistriodon are enlarged and point forwards rather than ventrally, and in this respect they are markedly different from those of other suids, including Libycochoerus massai . The skull of the latter species from Gebel Zelten, Libya, has small, vertically oriented bullae which are located far behind the pterygoid wings of the palatines.
If we add these characters to the others discussed by Pickford (1986, 1993), then it is difficult to escape the conclusion that kubanochoeres differ from listriodonts at least at the subfamily level. The differences in their skulls are as great or greater than those between the skulls of Listriodon and Sus which most authors readily accept as representing different subfamilies. Furthermore, these differences are present in the earliest members known of each of the subfamilies, from which it is concluded that the two subfamilies evolved from different ancestral groups.
If we include biogeographic evidence with the palaeontological data, then the separate evolution of the two groups becomes even more evident. Kubanochoeres evolved in Africa and then spread to the Middle East and Asia, but apparently never reached Europe. Listriodonts evolved in Eurasia (probably Europe) and then spread to Africa after they had become lophodont.
If we examine the palaeoecology of the two subfamilies, then we cannot fail to be struck by the fact that kubanochoeres evolved from small precursors the size of the pygmy hog ( Sus (Porcula) salvanius (Hodgson, 1847)) and that, with the passage of geological time, they experienced a regular and exaggerated increase in size, with the late middle Miocene species Megalochoerus khinzikebirus ( Wilkinson, 1976) being of gigantic size as large as a hippopotamus, whilst the largest known suid species ( Megalochoerus homungous Pickford, 1993 ) was as large as a gomphothere. Thus from about 20 Ma to 12 Ma the kubanochoeres followed a steep but classic Depéret-Cope body size trajectory during their evolution. Listriodonts, in strong contrast, started out being medium sized suids about the same size as Sus scrofa and remained virtually the same size throughout their history, except for the middle Miocene African genus Lopholistriodon which was smaller than Listriodon . Even the largest listriodonts, L. splendens major , of which the Toril cranium is a particularly large representative, is barely larger than that of a big wild boar. The main evolutionary trend in listriodonts was the development of lophodont cheek dentition and extremely wide incisors, and in this the subfamily converged functionally on the tapirs (folivorous perissodactyls) ( Leinders 1978). Kubanochoeres, in contrast, retained a bunodont cheek dentition, the largest species converging dentally to some extent with bunodont proboscideans. The middle Miocene species of this subfamily even developed cranial appendages on the frontal bone, the only group of suids known to do so.
Thus, morphological, palaeontological, biogeographic and palaeoecological evidence all point to the same conclusion – that Listriodontinae and Kubanochoerinae evolved from different ancestors on different continents and thereafter had quite separate evolutionary histories ( Pickford 1986). For all these reasons, it is concluded that the two groups should not be classed within the same subfamily, and we therefore recognise two subfamilies, Listriodontinae and Kubanochoerinae .
THE STATUS OF BUNOLISTRIODON
It has previously been argued ( Leinders 1975; Pickford 1986; Pickford & Moya Sola 1995) that the genus Bunolistriodon was erected in an invalid manner and that in any case the type and other European species assigned to it did not differ enough from Listriodon splendens to warrant separation at the generic level. Van der Made (1996, 1997b) has pointed out that even though the erection of the generic name was done in a most peculiar way, it is nevertheless valid according to the International Code of Zoological Nomenclature ( ICZN 1999).
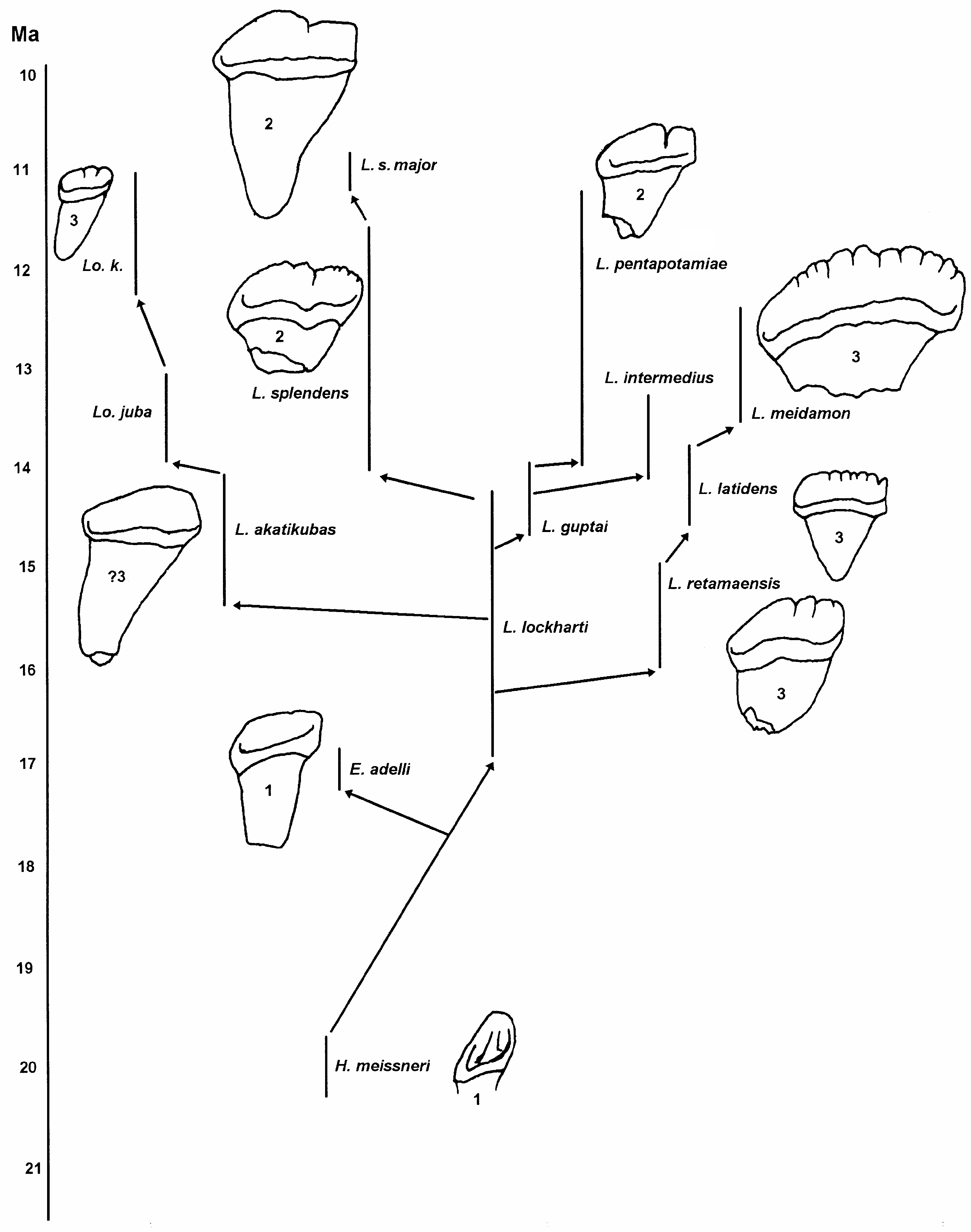
Re-examination of the fossils assigned to Bunolistriodon lockharti (the type species of Bunolistriodon ) and Listriodon splendens (the type species of Listriodon ) reveals that they are so similar in cranial, mandibular and postcranial anatomy that they are most probably congeneric, in which case Listriodon takes precedence over Bunolistriodon . The only significant differences between these two species relate to the degree of lophodonty of the cheek teeth and the anteroposterior lengthening of the incisors. With the passage of time the bunodont cheek dentition of L. lockharti gradually became more lophodont until the fully lophodont condition observed in L. splendens was achieved. There is no marked break in the morphometric continuum, nor is there evidence of two immigration events into Europe, one for Bunolistriodon and one for Listriodon . Leinders (1975) studied the problem of the affinities of the bunodont listriodonts of Europe, and concluded that the differences between Listriodon lockharti and Listriodon splendens were relatively unimportant and could not justify the continued generic subdivision between the two species. We fully concur with him. Under the circumstances, the safest and most stable conclusion to reach is that Bunolistriodon is a synonym of Listriodon .
We have considered whether Bunolistriodon should be kept on in the interests of stability of nomenclature, but have concluded that far from promoting stability, its continued use will merely add to the confusion that already exists. One of the reasons for the confusion is that the characters cited by Arambourg (1963) as defining the genus do not occur in it, but rather in the genera Libycochoerus and Kubanochoerus , both of which belong to a different subfamily, Kubanochoerinae ( Gabunia 1960) . Because of this, not only Arambourg (1963), but also other scientists ( Wilkinson 1976; Van der Made 1996, 1997b) have employed the generic name Bunolistriodon for several species of suids which are not listriodonts.
Van der Made (1996) included within Bunolistriodon a heterogeneous suite of suids, several species of which we consider to be kubanochoeres while others are listriodonts. Bunolistriodon affinis ( Pilgrim, 1908) , based on a fragmentary maxilla with P4 and M1, from Bugti, Pakistan, could well be a kubanochoere ( Pickford 1988), but until better material is collected from the type locality doubt will persist about the status of this species. Bunolistriodon anchidens Van der Made, 1996 , from Rusinga, Kenya, is a kubanochoere ( Pickford 1986) and we consider it to be a junior synonym of Libycochoerus jeanneli ( Arambourg, 1933) as it does not differ from the latter species either metrically or morphologically. Most of the material assigned by Van der Made (1996) to Bunolistriodon aff. latidens is equivalent to Listriodon retamaensis n. sp. and not to Eurolistriodon adelli as thought by Van der Made (1996). Bunolistriodon meidamon Fortelius, Van der Made & Bernor, 1996 is similar in many features to Listriodon latidens ( Biedermann 1873) . There can be little doubt that these two species are closely related. They occur within the same relatively restricted geographic area (southeastern Europe) and time period (MN5-6) and share several derived features in the morphology of the incisors (greatly elongated mesiodistally and with a crenulated apical border), canines (greatly enlarged) and cheek dentition (bunolophodont) ( Figs 9 View FIG ; 10 View FIG ). In passing we should note that Van der Made (1996: pl. 18, figs 1, 2) gives the holotype of this species as a left upper canine (NSSW 113) and a right and left mandible (NSSW 99) of which the symphysis ( Van der Made 1996: pl. 18, fig. 3) is said to be a paratype. This is erroneous, the mandible with the symphysis is the holotype, while the upper canine is the paratype. Bunolistriodon lockharti ( Pomel, 1848) , Bunolistriodon intermedius (Liu & Lee, 1963) ( Liu & Lee 1963a; Ye et al. 1992) and Bunolistriodon akatikubas Wilkinson, 1976 ( Wilkinson 1976) are all listriodonts of the genus Listriodon . Van der Made’s (1996) hypodigm of Bunolistriodon guptai ( Pilgrim, 1926) is, in our opinion, a mixture of kubanochoere and listriodont fossils, but the holotype is a Listriodon . In any case, Pickford (1988) considered L. guptai to be a synonym of L. pentapotamiae ( Falconer, 1868) .
RELATIONSHIPS BETWEEN LISTRIODONTINAE AND KUBANOCHOERINAE
Van der Made (1996, 1997b) has argued that kubanochoeres represent a tribe within the subfamily Listriodontinae , whereas Pickford (1993) considered that they are a separate subfamily of Suidae . Van der Made (1996: text fig. 58) indicated that his tribe Kubanochoerini consisted of only a single genus Kubanochoerus which was derived with doubt from Bunolistriodon affinis or from a form more primitive than Bunolistriodon affinis .
Van der Made’s arrangement of his tribe Listriodontini recognises two genera, Bunolistriodon and Listriodon . Skulls of some of the species assigned by Van der Made to Bunolistriodon are now known. The species Eurolistriodon adelli possesses a skull with morphology that is close in several derived features to that of Listriodon splendens , the most derived species of the large Listriodontini . The snouts of Listriodon retamaensis n. sp. from Bézian and Retama are also typically listriodont, even if shorter than that of Eurolistriodon . The male snout of L. latidens from Inönü, Turkey, is also typical of Listriodontini , possessing a widely flaring canine flange, and the same morphology occurs in the Chinese sample of Listriodon ( Chen 1986) . In all these forms, the male snout has wide canine flanges and the Ushaped posterior nares are retired a long distance behind the level of the third upper molars.
In the African species assigned to Bunolistriodon by Van der Made, however, (i.e. B. anchidens , B. jeanneli ) the skulls and mandibles show no close resemblance to those of European Listriodontini . For example they do not have retired posterior nares (which are in any case V-shaped) among a host of other features enumerated by Pickford (1986). Similarly, none of the species assigned to Kubanochoerini by Van der Made possesses expanded canine flanges in male individuals, and the early species ( Kenyasus rusingensis Pickford, 1986 , Libycochoerus jeanneli , L. massai ) do not have retired posterior nares.
All this suggests that the Listriodontinae as conceptualised by Van der Made comprises at least two suid lineages which have no close phylogenetic relationships to each other. It is likely that the Kubanochoerini of Van der Made is a natural clade, but it should have attached to it the African species put by him, erroneosly in our view, into the genus Bunolistriodon . Pickford (1986) previously argued on the basis of cranial, mandibular and dental evidence that the African species Libycochoerus jeanneli is a kubanochoere and not a listriodont, and we see no compelling reason to change his conclusions about it. If the species Libycochoerus anchidens proves to be separate from L. jeanneli , which we doubt, then it too would join the Kubanochoerinae .
ORIGINS OF LISTRIODONTINAE
By the onset of MN4a European listriodonts already comprised two genera, Eurolistriodon and Listriodon , with significant differences in cranial and postcranial anatomy. It is therefore likely that the initial radiation of the subfamily occurred in MN3 or MN2, but thus far no convincing listriodont ancestor has been identified in these levels. For this reason some authors, including Van der Made (1996) have proposed that listriodonts evolved elsewhere and then migrated into Europe during the faunal turnover that heralded European land mammal zone MN4, a period which saw proboscideans, among other mammals, colonise Eurasia from Africa. The main problem with this suggestion is that listriodonts are unknown anywhere else in the world in deposits of MN3 and MN2.
In a general way it has been suggested that the listriodonts evolved from a species of Hyotherium such as H. meisneri von Meyer, 1850 or H. major (see Fig. 18 View FIG ), but there has never been much conviction in these proposals, even though out of the known suids from MN2 and MN3, the genus Hyotherium possesses dental morphology which is closest to that of early listriodonts. What is required to settle the matter is a detailed study of the skulls, because it is now evident that in suids in general and in listriodonts in particular, it was the skull that diversified more rapidly than the dentition. Thus early listriodonts will probably be identifiable as such on the basis of their skull morphology, whereas their dentitions will not be
I 1 I 2 I 3 P2 P3 P4 M1 M2 M3 i1 i2 i3 p2 p3 p 4 m 1 m 2 m 3
Length upper dentition Length lower dentition
FIG. 19. — Size variation (in mm) of the lengths of the upper (left) and lower (right) teeth of three subspecies of Listriodon splendens von Meyer, 1846 from MN6, MN7/8 and MN9, showing significant overlap in measurements. Symbols:, maximum and minimum measures; l, range of variation.
markedly different from those of hyotheres. No doubt this is why some isolated teeth of early Listriodon have on occasion been identified as Hyotherium (Golpe-Posse 1972; Chang 1974). Eurolistriodon adelli possesses a skull with all the classic listriodont morphology ( Pickford & Moya Sola 1995) yet its cheek dentition is primitive, differing from that of Hyotherium meisneri principally by its superior size (Figs 16; 17). Even the upper central incisors of E. adelli are weakly elongated, being intermediate in length/breadth index between those of H. meisneri and those of L. lockharti ( Fig. 18 View FIG ).
The dentition of Aureliachoerus Ginsburg, 1974 differs in several important respects from those of early listriodonts, and it is unlikely that this genus is closely related to them. The original hypodigm of Xenohyus venitor Ginsburg, 1980 , a suoid known from MN2 and MN3 of southwestern Europe, originally thought to be a suid ( Ginsburg 1980) but later transferred to the Old World Tayassuidae ( Pickford & Morales 1989) , contains a few teeth that may well be suid, and there remains the possibility that the origin of the listriodonts could be found in the taxon represented by these specimens. However, its fossil record is so fragmentary and poor that, for the time being, very little can be said about it.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Listriodon splendens von Meyer, 1846
| Pickford, Martin & Morales, Jorge 2003 |
Listriodontinae
| Van der Made 1996 |
Listriodontinae
| Van der Made 1996 |
Lopholistriodon kidogosana
| Pickford & Wilkinson 1975 |
Lopholistriodon
| Pickford & Wilkinson 1975 |
Libycochoerus massai
| Arambourg 1961 |
Libycochoerus massai
| Arambourg 1961 |
Listriodon
| von Meyer 1846 |
Listriodon
| von Meyer 1846 |
Listriodon
| von Meyer 1846 |