
Noblella coloma Guayasamin & Terán, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.188980 |
|
DOI |
https://doi.org/10.5281/zenodo.5627998 |
|
persistent identifier |
https://treatment.plazi.org/id/03871922-FFF9-ED2E-FF4A-FD26FE67FD51 |
|
treatment provided by |
Plazi |
|
scientific name |
Noblella coloma Guayasamin & Terán |
| status |
sp. nov. |
Noblella coloma Guayasamin & Terán , new species
Holotype. QCAZ 26307, adult male ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 ), collected from Reserva Florística Río Guajalito (0°14' S, 78°49' W; 1800–2000 m), 3 km NW from Km 59 on the old road from Quito to Santo Domingo, collected by Rubén D. Jarrín on 18 October 2003.
Paratopotypes. QCAZ 7277, adult female, collected by Leonora Orejuela on 22 Abril 1995; QCAZ 8701, juvenile female, collected by Santiago Burneo on 15 October 1995; QCAZ 11614, juvenile, collected by Ítalo G. Tapia, date unknown; QCAZ 32702, juvenile, collected by Cristina E. Toapanta on 16 December 2006; QCAZ 6466, sex unknown, collected by George Fletcher on 23 October 1994, cleared-and-stained; QCAZ 7412, sex unknown, collected by Juan M. Guayasamin on 18 June 1995, cleared-and-stained; QCAZ 40579, juvenile female, collected by Diana Pazmiño on 15 March 2009.
Generic placement. Duellman (1991), De la Riva et al. (2008), and Hedges et al. (2008) list the following traits as characteristic of Noblella : (1) head narrower than body; (2) tympanic membrane differentiated (except in N. duellmani ); (3) cranial crests absent; (4) dentigerous processes of vomers absent; (5) “S” condition of adductor muscle; (6) terminal discs on fingers and toes not expanded or slightly expanded; discs and circumferential grooves present distally (except in N. duellmani ); terminal phalanges narrowly T- shaped; (7) Finger I shorter than, or equal in length to, Finger II; (8) Toe III shorter than Toe V; tips of at least Toes III–IV pointed; (9) subarticular tubercles not protruding; (10) conspicuous tarsal tubercle; (11) dark inguinal spots present (except in N. duellmani ); (12) small size (SVL <22 mm). All the traits observed in the new species agree with the diagnosis of Noblella ; therefore, the placement of Noblella coloma in the genus Noblella is justified. The only other possibility for the generic placement of the new species is Barycholos , a genus endemic to South America that has a close evolutionary and morphological affinity with Noblella (Heinicke et al. 2007; Hedges et al. 2008). However, species in Barycholos have vomers with small dentigerous processes (absent in Noblella ), Finger I longer than II (Finger I shorter than, or equal in length to, Finger II in Noblella ), and projecting subarticular tubercles (not projecting in Noblella ).
Diagnosis. Noblella coloma ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 ) presents the following characteristics: (1) skin of the dorsum finely shagreen; (2) tympanic annulus and membrane clearly visible and differentiated, supratympanic fold covering upper border of tympanum ( Fig. 2 View FIGURE 2 C); (3) snout rounded in dorsal and lateral view (eye to nostril distance 48.8–58.9% of eye diameter, Fig. 2 View FIGURE 2 C); (4) dentigerous processes of vomers absent; (5) fingers not expanded distally; finger tips acuminate; Finger I slightly shorter than Finger II ( Fig. 3 View FIGURE 3 ); nuptial pad not visible; fingers lacking circumferential grooves; (6) distal phalanges T-shaped; phalangeal formula of hands: 2, 2, 3, 3 ( Fig. 5 View FIGURE 5 ); (7) hands with one weakly defined supernumerary tubercle; subarticular tubercles rounded, not prominent; ulnar tubercles absent; (8) one elongated tarsal tubercle ( Fig. 3 View FIGURE 3 ); two prominent metatarsal tubercles; supernumerary plantar tubercles absent; toes slightly expanded and acuminate distally; (9) Toe V shorter than Toe III; distal portions of circumferential grooves present on Toes III–V; (10) phalangeal formula of feet: 2, 2, 3, 4, 3 ( Fig. 5 View FIGURE 5 ); (11) in life, dorsum uniform brown, except for distinctive suprainguinal marks; flanks with dark brown band that narrows as it approaches groin; venter orange with minute brown and white spots ( Fig. 1 View FIGURE 1 ); (12) in one adult male, SVL 14.6 mm; in one adult female, 16.0 mm.
Noblella coloma differs from all the other species in Noblella by lacking most of the dorsal color marks (i.e., interobital bar, scapular and sacral chevrons; Fig. 2 View FIGURE 2 ), and by having a bright orange venter ( Fig. 1 View FIGURE 1 ). Additionally, the dorsum of N. coloma is shagreen, whereas the dorsum of N. duellmani and N. lynchi is pustular, and N. pygmaea has a tubercular dorsal skin. Noblella coloma is similar to N. duellmani , N. heyeri , N. lynchi and N. pygmaea in having three phalanges in Finger IV, in contrast with N. carrascoicola , N. lochites , N. myrmecoides , and N. ritarasquinae that have two phalanges in Finger IV. A tympanum is evident in N. coloma , N. heyeri , N. lynchi , N. myrmecoides , N. peruviana , and N. pygmaea , and is absent in N.
duellmani , N. carrascoicola , and N. ritarasquinae . In preservative, N. coloma has a dark throat and a uniform cream venter, whereas the venter is brown with cream mottling in N. duellmani , N lynchi , N. heyeri , and N. carrascoicola ( Fig. 6 View FIGURE 6 ). Comparison of the new species with Noblella peruviana are difficult because of the paucity of information regarding the morphology of the later. All what we know about N. peruviana comes from the brief description provided by Noble (1921) and De la Riva et al. (2008), both based on five specimens collected more than a century ago. Characters mentioned in Noble’s original description include: tympanum present, digits pointed with no terminal disks, well-developed and elongated tarsal tubercle, skin feebly granular above and smooth below, dorsal coloration uniform gray brown or with dark brown blotches, and broad band of dark brown extending on each side from the nostril to the lumbar region. Additionally, De la Riva et al. (2008) mentioned that the types of N. peruviana have the tips on Toes II, III and IV distinctively pointed, and that, at least in some paratypes, a black inguinal spot is present. The traits listed above for N. peruviana are present in several other species of the genus; therefore, comparisons are limited. However, Noble (1921) mentioned that, in preservative, the ventral coloration in N. peruviana was reddish brown; in contrast, the ventral coloration of N. coloma is cream. In addition, N. coloma and N. heyeri are the only species in the genus found on the western slopes of the Andes ( Table 1 View TABLE 1 ); this being a biogeographical criterion that could be useful to distinguish among species.
Description of holotype. Adult male (QCAZ 26307); head as long as wide; snout round in dorsal and lateral views ( Fig. 2 View FIGURE 2 C); canthus rostralis straight; loreal region concave; upper eyelid about 50% of interorbital distance; eye-nostril distance 59% of eye diameter; tympanum visible almost completely, tympanic membrane well differentiated from surrounding skin; supratympanic fold covering upper border of tympanum. Dentigerous processes of vomers absent. Skin of dorsum finely shagreen; venter smooth; ulnar tubercles absent; palmar tubercle oval, about 1.5x the size of thenar tubercle; one supernumerary tubercle weakly marked; proximal subarticular tubercles large, rounded; fingers not expanded distally, finger tips slightly acuminate ( Fig. 3 View FIGURE 3 ), circumferential grooves absent; relative lengths of fingers I <II <IV <V.
One prominent tarsal tubercle on ventral surface of tarsus ( Fig. 3 View FIGURE 3 ); metatarsal tubercles conspicuous, similar size, longer than broad; proximal subarticular tubercles visible, distal subarticular tubercles inconspicuous; no supernumerary plantar tubercles. Toes slightly expanded and acuminate distally; distal portions of circumferential grooves present on Toes III–V; relative lengths of toes I <II <V <III <IV. For measurements of the type series (mm) see Table 2 View TABLE 2 .
heyeri Ecuador, Peru: Andes of northern Peru (33 km SW of 2000–3100 m Lynch (1986), this work Huancabamba) and southern Ecuador (Provincia de Loja).
Color of holotype in alcohol. Dorsum grayish brown to brown with dark brown inguinal spots; scapular and sacral chevrons absent. Sides of head uniform black continuous posteriorly onto flanks; labial bars absent; each flank with a wide longitudinal dark brown stripe that narrows posteriorly. Forearms and hind limbs with well-defined dark brown marks ( Fig. 2 View FIGURE 2 ). Throat dark brown; venter cream with minute dark spots (visible under magnification).
Coloration in life. Dorsum orange-brown, with a thin dark brown middorsal stripe and dark brown inguinal spots; interobital bar and scapular and sacral marks absent. Venter orange with minute white and brown spots ( Fig. 1 View FIGURE 1 ). Sides of head dark brown; flanks with dark brown stripe that narrows posteriorly. Iris dark brown with orange ring around pupil (based on photographs of QCAZ 32702, holotype, by Luis A. Coloma ).
Variation. Throat cream (QCAZ 32702) or cream with minute brown spots visible under magnification (QCAZ 11614). There is no evident sexual dimorphism. For measurements see Table 2 View TABLE 2 .
7277 26307 8701 40579 11614 32702 Sex Adult female Adult Male Juv. Female Juv. female Juvenile Juvenile Osteology. The following osteological description of Noblella coloma is based on a cleared-and-double stained individual (QCAZ 6466, SVL = 14.1 approximately; Figs. 4 View FIGURE 4 , 5 View FIGURE 5 ). The specimen lacks information regarding its sex, but because of its SVL we assume that it could be an adult or subadult male or juvenile female. Drawings of hand and foot are based on QCAZ 7412.
Cranium. Shape and proportions. The skull is widest posterior to the orbit at the level of the articulation of the maxilla with the quadratojugal ( Fig. 4 View FIGURE 4 ); at this level, the skull is as wide as long. The rostrum is short, accounting for less than a quarter (17%) of the medial length of the skull. At the level of midorbit, the width of the braincase is about 35% of the greatest width of the skull and 38% of the medial skull length.
Neurocranium. The anterior neurocranium comprises large olfactory capsules, which are cartilaginous, and the anterior part of the braincase. The medial walls of the nasal capsules are narrowly separated, with the internasal septum apparently being composed of a thin plate of cartilage between the capsule walls; a minute, ossified septomaxilla is present, but obscured by the staining of the anterior nasal capsule cartilages ( Fig. 4 View FIGURE 4 ).
The cartilaginous planum antorbitale has a slight anterolateral orientation in dorsal and ventral aspects. Ventrally, the planum is invested by a simple, thin and relatively long neopalatine ( Fig. 4 View FIGURE 4 ). The distal end of the neopalatine is clearly separated from the maxilla; the medial end articulates with the sphenethmoid ( Fig. 4 View FIGURE 4 ).
The braincase and otic capsules are moderately ossified. Ventrally, the bony sphenethmoid is not fused medially ( Fig. 4 View FIGURE 4 ). There is a moderately broad separation between the sphenethmoid and prootics, within which the optic fenestra lies ( Fig. 4 View FIGURE 4 ). The optic fenestra lies in cartilage, except for its posterior margin that is formed by the prootic bone. Dorsally and ventrally, the exoccipitals are separated from each other and from the prootics ( Fig. 4 View FIGURE 4 ). The contralateral prootics are completely separated from each other; each one bears a relatively long prootic foramen, and two small foramina, the oculomotor and trochlear. The frontoparietal fontanelle is partially exposed medially between the frontoparietal ( Fig. 4 View FIGURE 4 ).
Dorsally, the otic capsule is well ossified, and bears a moderately broad, cartilaginous crista parotica. The epiotic eminences are prominent; the anterior eminence is slightly longer than the posterior and the angle between the two arms is approximately 90˚. Ventrally, the lateral wall of the otic capsule is cartilaginous. The cartilaginous operculum lies posteriorly adjacent to the pars interna plectri of the stapes in the fenestra ovalis, which is formed entirely in cartilage. The pars media plectri is a long, slender, ossified element that extends anterolaterally beneath the crista parotica and behind the palatoquadrate and ventral ramus of the squamosal to terminate in the club-shaped cartilaginous pars externa plectri located in the middle of the incomplete tympanic annulus ( Fig. 4 View FIGURE 4 ).
Dorsal investing bones. Dorsal investing bones are poorly developed. The nasals are thin and widely separated from one another; they cover a large area of the nasal capsules dorsally. The frontoparietals are well-developed bones, separated medially, and separated from the prootics. The frontoparietal bears a lamina perpendicularis, the vertical component forming the dorsolateral edge of the braincase ( Fig.4 View FIGURE 4 ).
Ventral investing and palatal bones. Owing to the thinness of the bone, the posterior margin of the parasphenoid is difficult to determine accurately. The cultriform process (ca. 39% the width of the braincase) extends anteriorly reaching about the midlength of the sphenethmoid ( Fig. 4 View FIGURE 4 ). The anterior end of the process is tapered; the parasphenoid is separated from the neopalatines. The lateral margins of the process are approximately parallel; anteriorly, the margins converge gradually towards one another and make contact with the medial margin of the sphenoid. The parasphenoid alae are moderately long (about equal to the width of the cultriform process), slightly posterolaterally oriented, and distally truncate beneath the midwidth of the otic capsules. A posteromedial process is present, but distinctly separated from the margin of the foramen magnum ( Fig. 4 View FIGURE 4 ).
The small vomers are broadly separated from one another medially. Each is composed of an arcuate bone bordering the medial and small portions of the anterior and posterior margins of the choana. The prechoanal and postchoanal ramus are slightly expanded anteriorly; the dentigerous process is absent ( Fig. 4 View FIGURE 4 ).
The neopalatines are thin and underlie the plana antorbitalae, covering the posterior surfaces and dorsomedial surfaces of these structures ( Fig. 4 View FIGURE 4 ). The bones are unornamented, straight, and articulate with the sphenethmoid; each neopalatine is separated from the maxilla.
Maxillary arcade. The maxillary arcade bears teeth on the premaxillae and maxillae. The maxillae are in close proximity to the quadratojugals, but have no contact ( Fig. 4 View FIGURE 4 ). The partes palatinae of the maxillae are narrow and barely evident. The premaxilla bears prominent partes palatinae; the posterolateral palatinae process is more robust than the posteromedial process ( Fig. 4 View FIGURE 4 ). There is a simple, juxtaposed articulation between the anterior end of the maxilla and the premaxilla. The pars facialis of the maxilla is well developed anteriorly, having a preorbital process that covers the planum antorbitale along the lateral aspect of the olfactory capsule ( Fig. 4 View FIGURE 4 ).
Suspensory apparatus. The triradiate pterygoid bears a curved anterior ramus that is oriented anterolaterally toward the maxilla, with which it has an area of close proximity. The pterygoid is separated from the maxilla by the pterygoid cartilage, which lies along the medial margin of the maxilla in the orbital region. The medial and posterior rami of the pterygoid are about equal in length; however, the medial ramus is more robust than the posterior. The edge of the medial ramus overlaps the lateral edge of the ossified margin of the prootic.
The squamosal is T-shaped; the otic ramus is much more longer than the zygomatic ramus (otic ramus 3 x the length of the zigomatic ramus). The otic ramus overlaps a large area of the crista parotica. The ventral ramus invests the lateral surface of the palatoquadrate, and articulates with the quadratojugal ( Fig. 4 View FIGURE 4 ).
Postcranium
Vertebral column. There are eight presacral vertebrae ( Fig. 5 View FIGURE 5 ). Presacral I is notably shorter than the posterior presacrals, and Presacral III is relatively longer than the other presacral vertebrae. All of the presacrals are non-imbricate. The vertebral profile in increasing order of overall width is: I <sacrum <VIII <VII = VI <V = II <IV <III.
Presacral I lacks a transverse process. The transverse processes of Presacral II have a slightly anterolateral orientation, Presacrals III–VI have a posterolateral orientation and Presacrals VII and VIII have an anterolateral orientation. The bony sacral diapophysis is not broadly expanded, and its width is about the same along all the bone. The leading edge of the diapophyses is straight and slightly orientated posterolaterally, whereas the posterior margin is concave, and also oriented posterolaterally. The urostyle is long, slender, and about equal length of the presacral portion of the vertebral column. The bone has a bicondylar articulation with the sacrum, and bears a prominent dorsal crest throughout most of its length ( Fig. 5 View FIGURE 5 ).
Pectoral girdle. The prezonal portion is represented by a short cartilaginous omosternum. The clavicles are oriented anteromedially, with the medial tips distinctly separated from one another and located at about the same level of the anterolateral end of the clavicle that articulates with the scapula. The coracoid is stout, with the glenoidal and sternal ends about equally expanded. The midshaft width is about 13% the length of the coracoid, and about 41% the width of the expansion of the sternal end of the bone. The pectoral fenestra is oval and about two times as wide as it is deep. The scapula is long with a prominent pars acromialis that is not separated from the pars glenoidalis. The clavicle is 75% the length of the scapula. The suprascapula is mineralized, with the ossified cleithrum apparent as a slender bone along the leading edge of the suprascapular blade and with an expansion of its proximal end. The sternum is mineralized and narrowly separated from the epicoracoid ( Fig. 5 View FIGURE 5 ).
Pelvic girdle. The long, slender ilial shafts bear conspicuous dorsolateral crests. The ilial prominence is broad and low; the pubes is cartilaginous. The round acetabulum is ossified ( Fig. 5 View FIGURE 5 ).
Manus and pes. The phalangeal formulae for the hand and foot are standard: 2-2-3-3 and 2-2-3-4-3, respectively. In increasing order of length, the order of the digits on the hand is: I <IV <II <III, and that of the foot is: I-II-V-III-IV. The carpus is composed of a radiale, ulnare, Element Y, Carpal 1, and a large postaxial element assumed to represent a fusion of Carpals 2, 3, and 4 ( Fig. 5 View FIGURE 5 ). The prepollex is composed of one small, proximal bone and an elongated distal cartilage. The terminal phalanges are T-shaped. The tarsus is composed of two tarsal elements, presumably Tarsal 1 and Tarsal 2 + 3. In a ventral view, three sesamoids (of about equal size) are covering the proximal end of the metatarsals III–IV. The prehallux is represented by a bony, rounded basal element associated with a distal cartilaginous element ( Fig. 5 View FIGURE 5 ).
Etymology. The specific name coloma is a patronym for Luis Aurelio Coloma in recognition of his continual efforts in studying and protecting amphibians, and mentoring students (us included). The specific epithet is used as a substantive in apposition.
Distribution and conservation. Noblella coloma is known only from the type locality, Reserva Florística Río Guajalito (0°14' S, 78°49' W; 1800–2000 m), 3 km NW, by path, from Km 59 on the Quito-Chiriboga- Santo Domingo road ( Fig. 7 View FIGURE 7 ). All individuals of N. coloma have been found active on leave litter during the day. The type locality of Noblella coloma is included in the Bosque de Neblina Montano (Montane Cloud Forest) according to the classification proposed by Valencia et al. (1999). Noblella coloma only is known from the type locality, within a private reserve. During the last 15 years, several groups of biology students have been visiting this reserve. As a result, only eight specimens have been collected, suggesting that the species is rare and/or very difficult to locate. Following the IUCN (2001) criteria, we consider N. coloma as data deficient.
Remarks. Species of Noblella share several morphological characteristics, thereby making this group easy to identify and, likely, monophyletic. Potentially derived traits present in most species of Noblella include pointed toes and conspicuous tarsal tubercle ( Fig. 3 View FIGURE 3 ). The thin neopalaties, reduced vomers, and lack of fusion among several bones (sphenethmoid, exoccipitals, prootics) could also be derived traits related to the small size of Noblella .
Given that all species except the Amazonian N. myrmecoides are endemic to the Andes ( Table 1 View TABLE 1 ), the most likely scenario is that this group originated in the Andes and later dispersed to the Amazon basin. The recent phylogeny of Hedges et al. (2008) places the only sampled species of Noblella ( N. lochites ) as the closest relative of Barycholos .


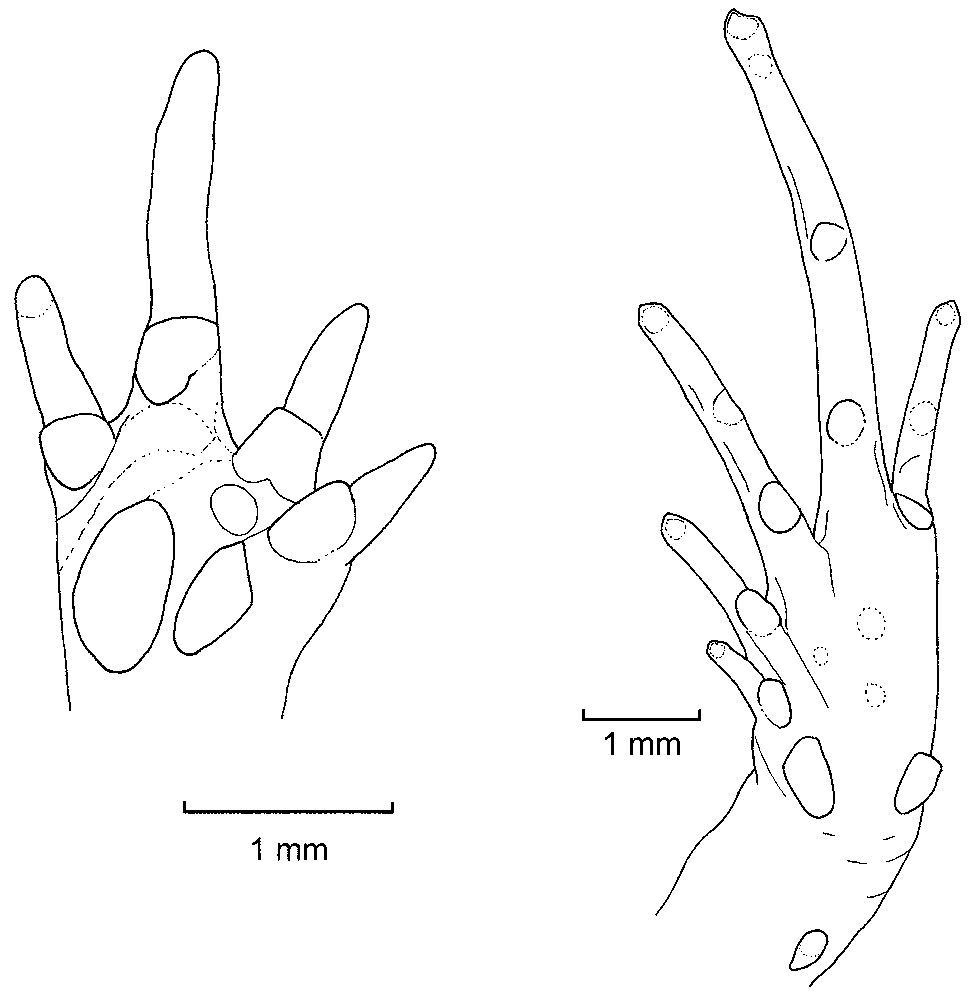
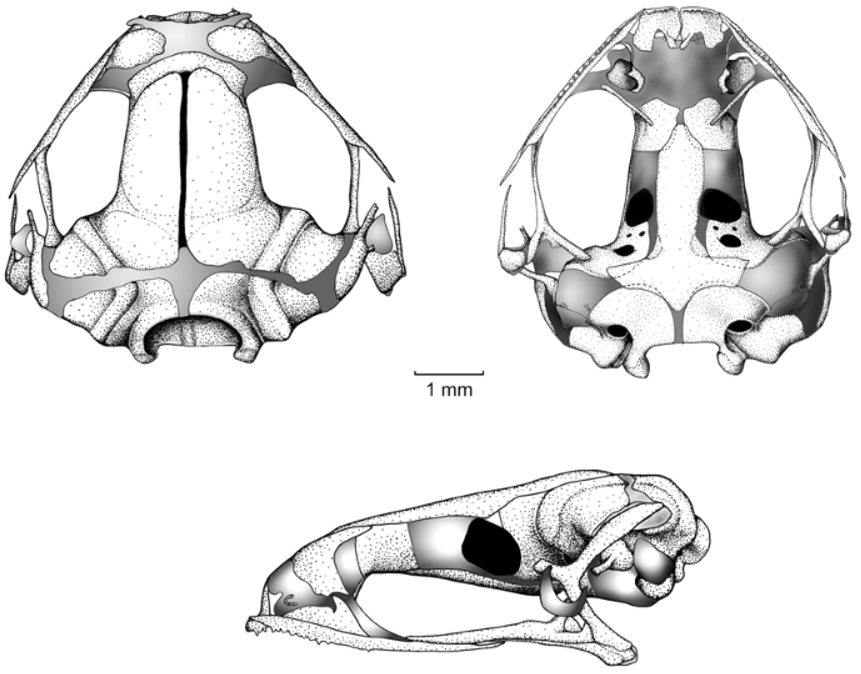
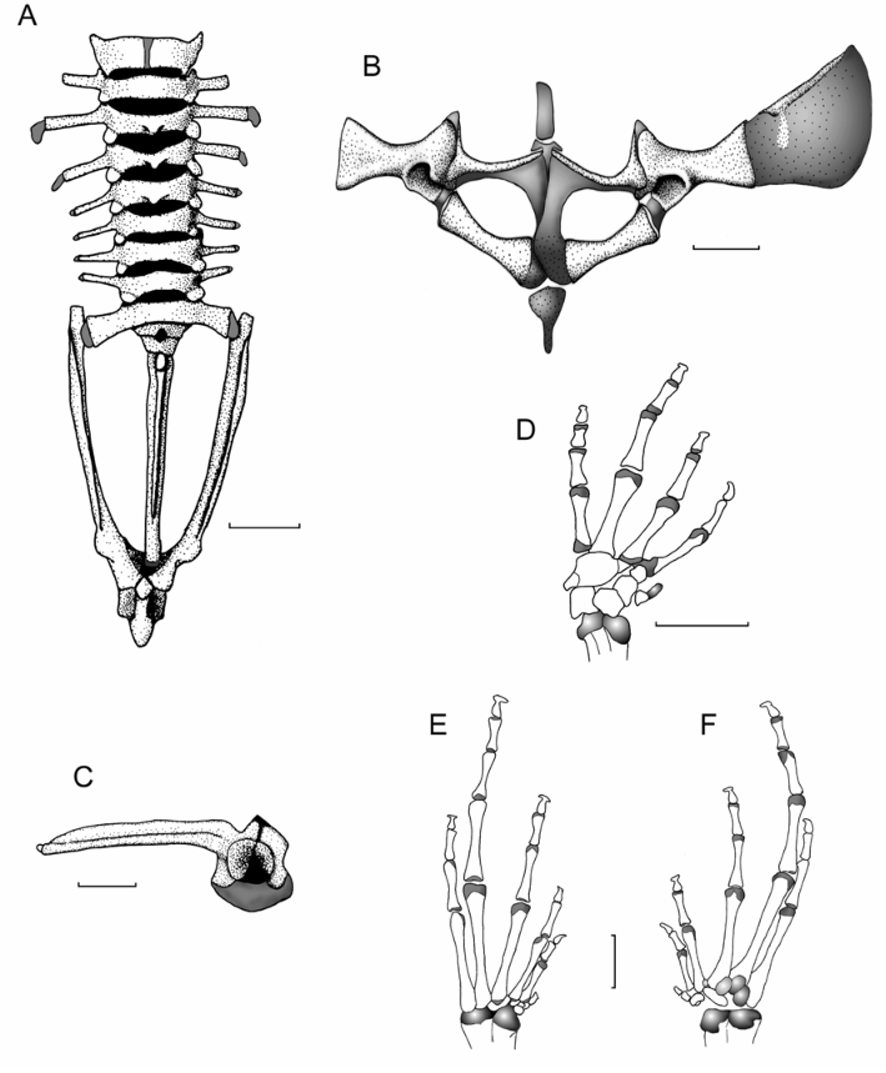

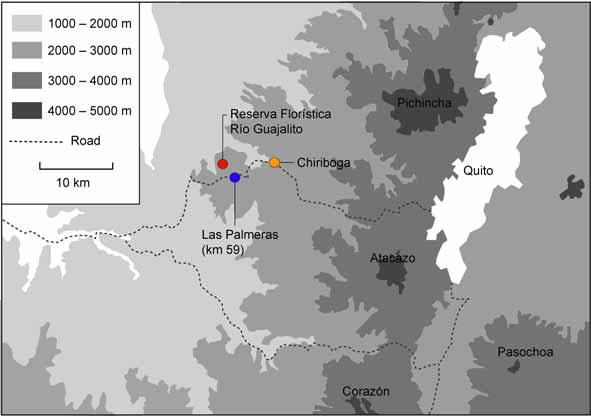
TABLE 1. Distribution of species in the genus Noblella.
| Species | Distribution | Elevation | Source |
|---|---|---|---|
| carrascoicola | Bolivia: Cloud forests along the northeastern Andean slopes from Chapare, Departamento Cochabamba, eastward to Provincia Caballero, Departamento Santa Cruz. | 1850–2700 m | De la Riva and Köhler (1998); Köhler (2000) |
| coloma | Ecuador: Western slope of the Andes, Provincia Pichincha, Reserva Florística Río Guajalito (0°14' S, 78°49' W). | 1800–2000 m | This work |
| duellmani | Peru: Eastern slope of the Andes, Departamento Pasco, Distrito Paucartambo, Cillapata (approximately 1.5 km NNE of Auquimarca; 10˚43'52" S, 75˚42'48" W). | 2900 m | Lehr et al. (2004) |
TABLE 2. Measurements of the type series (in mm) of Noblella coloma.
| SVL 16.03 | 14.55 | 12.61 | 11.28 | 10.26 | 10.65 |
|---|---|---|---|---|---|
| Tibia length 7.23 | 6.73 | 6.18 | 6.13 | 5.09 | 5.69 |
| Foot length 7.30 | 6.31 | 6.10 | 5.78 | 4.36 | 5.46 |
| Head length 5.09 | 4.51 | 4.29 | 4.17 | 3.44 | 3.66 |
| Head width 5.01 | 4.31 | 4.21 | 4.04 | 3.46 | 3.58 |
| Interorbital distance 2.01 | 1.71 | 1.82 | 1.60 | 1.50 | 1.46 |
| Upper eyelid width 1.04 | 0.86 | 1.07 | 0.91 | 0.89 | 0.88 |
| Internarial distance 2.08 | 1.80 | 1.76 | 1.57 | 1.42 | 1.38 |
| Eye to nostril distance 1.15 | 1.02 | 0.87 | 0.81 | 0.72 | 0.67 |
| Snout to eye distance 2.10 | 1.92 | 1.95 | 1.79 | 1.50 | 1.61 |
| Eye diameter 1.96 | 1.72 | 1.75 | 1.61 | 1.32 | 1.37 |
| Tympanum diameter 1.08 | 0.75 | 0.58 | 0.66 | 0.42 | 0.58 |
| Eye to tympanum distance 0.47 | 0.46 | 0.31 | 0.34 | 0.50 | 0.41 |
| Forearm length 3.63 | 3.63 | 3.17 | 2.76 | 2.48 | 2.79 |
| Hand length 2.59 | 2.89 | 2.37 | 2.62 | 2.44 | 2.48 |
| Finger I length 1.76 | 1.96 | 1.66 | 1.64 | 1.64 | 1.77 |
| Finger II length 1.80 | 2.11 | 1.76 | 1.73 | 1.76 | 1.86 |
| QCAZ |
Museo de Zoologia, Pontificia Universidad Catolica del Ecuador |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.