
Megamphicyon carnutense ( ANTUNES et GINSBURG, 1977)
|
publication ID |
https://doi.org/ 10.37520/fi.2021.011 |
|
persistent identifier |
https://treatment.plazi.org/id/0385BC16-FFF5-FFDB-FCEA-FDA0FA5C48E1 |
|
treatment provided by |
Felipe |
|
scientific name |
Megamphicyon carnutense ( ANTUNES et GINSBURG, 1977) |
| status |
|
Megamphicyon carnutense ( ANTUNES et GINSBURG, 1977)
Text-figs 5 View Text-fig , 6 View Text-fig Tabs 1b, 2b
1965 Pseudocyon sansaniensis aff. sansaniensis LARTET 1851; Kuss, pp. 119–122, fig. 77.
1977 Amphicyon giganteus carnutense ; Antunes and Ginsburg, p. 341.
1989 Amphicyon giganteus carnutense Antunes et Ginsburg, 1977 ; Ginsburg, p. 102, figs 1–4.
2000b Amphicyon (Megamphicyon) lathanicus n. sp.; Ginsburg et al., p. 607, fig. 6.
2003 Megamphicyon giganteus ; Fejfar et al., p. 167.
2016 Megamphicyon ‘ major – giganteus ’; Fejfar and Heizmann, p. 320, figs 7.2, 12.
2019 Amphicyon carnutense ; Jiangzuo et al., p. 2.
H o l o t y p e. NMB S.O. 6531, left mandible with canine and p4–m2.
T y p e l o c a l i t y. Chilleurs, France.
A g e. Early Miocene, MN 3.
D i a g n o s i s. In Antunes and Ginsburg (1977).
S t u d i e d m a t e r i a l f r o m T u c h o ř i c e. NMPv 11701 (TU 738911), right P4; NM-Pv 11703 (TU 738910), P4 left; NM-Pv 11704 (TU 73892), left M1; NM-Pv 11705 (TU 738912), left M2; NM-Pv 11706 (TU 738916), left M3; NM-Pv 11707 (TU 738920), left M3; NM-Pv 11708 (TU 73898), left p4; NM-Pv 11709 (TU 738919b), left p4; NMPv 11747 (TU 739152), left m1–m3; NM-Pv 11710 (TU 738918), left m1; NM-Pv 11711 (TU 73896), left m1; NMPv 11712 (TU 739154), left m1–m2; NM-Pv 11713 (TU 73914), left m2; NM-Pv 11714 (TU 73893), left m2; NMPv 11716 (TU 73897), left m2; NM-Pv 11717 (TU 73899), left m2; NM-Pv 11718 (TU 738913), right m2; NMPv 11696 (TU 739120/21), association of right m2 and m3; NM-Pv 11719 (TU 738914a), left m3.
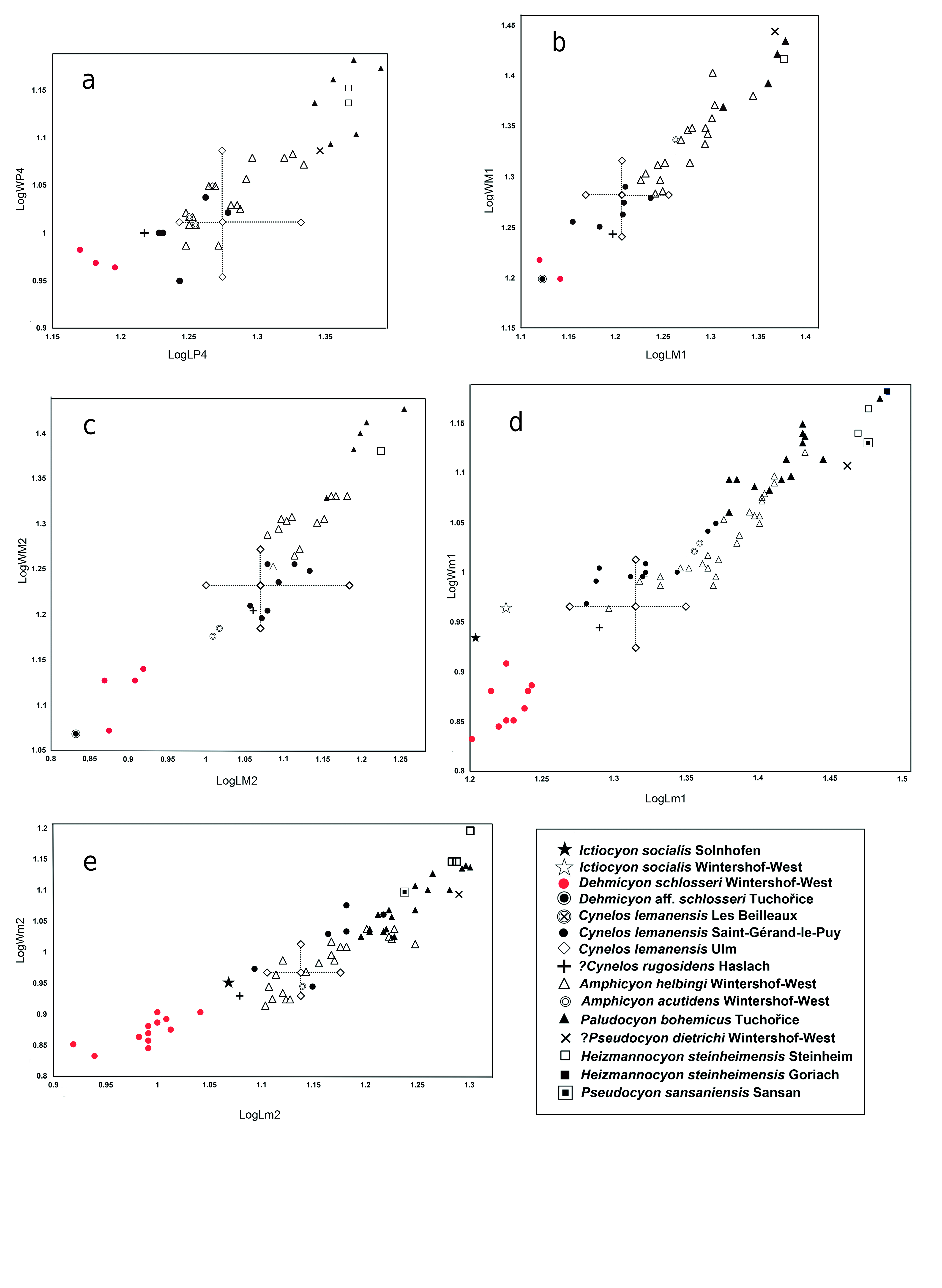
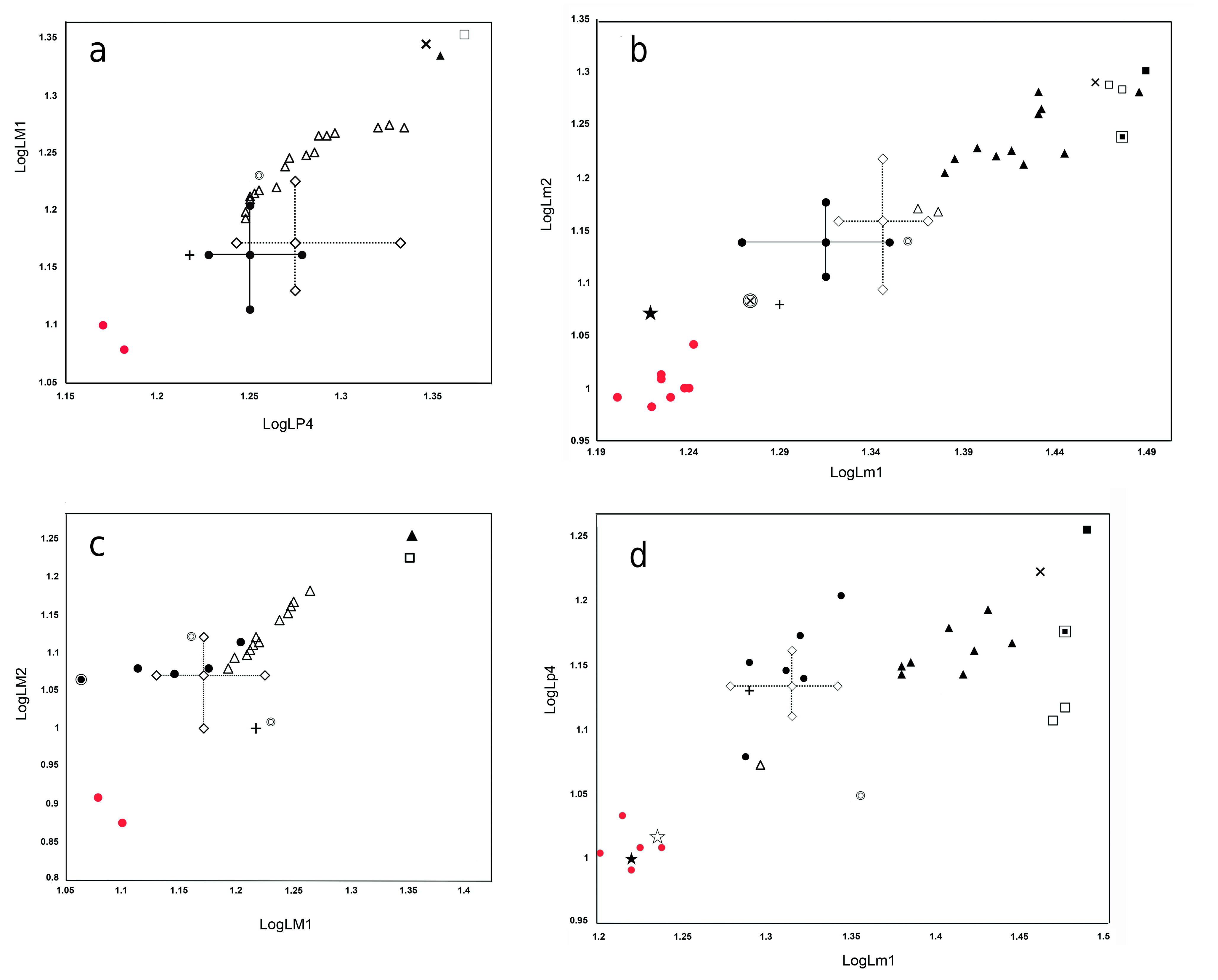
R e m a r k s. The large-sized Miocene amphicyonids of Western Europe have frequently been determined as Amphicyon giganteus ( SCHINZ, 1825) or Megamphicyon giganteus ( SCHINZ, 1825) , depending on the authors ( Kuss 1965, Ginsburg and Antunes 1968, Ginsburg 1999, Peigné et al. 2006). Some of the large species from the early Miocene have been classified in other amphicyonid genera such as Ysengrinia and Crassidia ( Heizmann and Kordikova 2000) and even in Pseudocyon ( Ginsburg 1967, 1999, Heizmann and Kordikova 2000). These three genera exhibit different degrees of hypercarnivorous dental adaptation ( Morales et al. 2019), which serves to separate them from the Amphicyon major group, which reveals a tendency to enlarge the crushing molar surface.
From at least the late Oligocene to the middle Miocene, a set of large forms is recorded in Europe, which are close in size and morphology to Amphicyon major, but show some differences which are very difficult to evaluate ( Kuss 1965). Increased dental size can be recognised when comparing from the oldest form to the most modern ones, but due to the great variability in size and morphology, there are real difficulties involved in assigning these fossil assemblages to different taxa. Some of the older forms from the early Miocene (MN 2–3) have received different determinations, including Amphicyon giganteus carnutense ANTUNES et GINSBURG 1977 , Amphicyon giganteus laugnacensis GINSBURG, 1989 and Amphicyon (Megamphicyon) lathanicus GINSBURG, CHENEVAL, JANVIER, POUIT et SEN, 2000 (see Ginsburg 2000b, 2002). However, the type species, Megamphicyon giganteus ( SCHINZ, 1825) , would be reserved for the largest species with molars similar in size to those from the type locality of Averay ( France), also found in some European localities of MN 4–6 age (Artenay, Baigneaux, La Romieu, Pont Levoy and Arroyo del Val among others), where molar sizes can greatly exceed that of Amphicyon major ( Ginsburg 1989, 1999, Peigné et al. 2006). However, there is a need for an indepth review of this group, a task that lies beyond the scope of the present research.
D e s c r i p t i o n. NM-Pv 11701, right P4 ( Text-fig. 5a View Text-fig 1–3 View Text-fig View Text-fig View Text-fig ). Quite elongated, with a relatively long metastyle, and a narrow mesially widened paracone, although without developing an authentic parastyle. The mesial crista of the paracone is strogly pronounced, with a small incision near the base, which changes its inclination, tending to become more horizontal and displaced buccally. Consequently, the base of the paracone is continued mesially. The protocone is moderate in size, placed in the mesial position, with a poorly separated cusp. Highly developed basal cingulum.
NM-Pv 11703, P4 left.This tooth presents an intermediate size, somewhere between the two previous specimens; the buccal part of the metastyle is broken. The anterior crista of the paracone is clearly more vertical than in the other two specimens.
NM-Pv 11704, left M1 ( Text-fig. 5b View Text-fig 1–2 View Text-fig View Text-fig ). Molar with a subtriangular occlusal shape, the base of the lingual wall is somewhat broken; whether a basal cingulum existed is not evident. The paracone is robust and large, the metacone is low and somewhat smaller. Buccal styles scarcely developed. The large pyramidal protocone does not reveal the presence of a paraconule, but a rather large metaconule can be observed. Cingulum buccal very strong.
NM-Pv 11705, left M2 ( Text-fig. 5c View Text-fig ). Molar quite wide, but narrow. Paracone very pronounced in relation to the metacone, which is somewhat displaced lingually. The protocone and the lingual cingulum are well developed.
NM-Pv 11706, left M3 ( Text-fig. 5d View Text-fig ), compared to the previous molar it is smaller, but the height and strength of its buccal cones rule out any possible interpretation as an M3, although there is a strong constriction in the distal wall, which is similar to that observed in the specimen NMPv 11707, which is considered as an M3.
NM-Pv 11707, left M3 ( Text-fig. 5e View Text-fig ). Similar in morphology to the M3 described above, but with very poorly developed cusps. The metacone is very small, almost lost. Occlusal kidney-shaped form. Very strong buccal cingulum, which represents almost the most developed element of the molar.
NM-Pv 11708, left p4 ( Text-fig. 6a View Text-fig 1–3 View Text-fig View Text-fig View Text-fig ). Premolar with strong and high distal cuspid. Distolingual cingulid well developed and talonid moderately expanded.
NM-Pv 11709, left p4 ( Text-fig. 6b View Text-fig 1–3 View Text-fig View Text-fig View Text-fig ), smaller than the previous specimen and differs from it in the strong constriction of the lingual wall.
NM-Pv 11710, left m1 ( Text-fig. 6c View Text-fig 1–3 View Text-fig View Text-fig View Text-fig ). Very robust molar, with a short paraconid and the mesial cristid distally inclined. High and very robust protoconid. Metaconid quite reduced, slightly displaced distally and with a somewhat swollen buccal wall. Short talonid, almost completely occupied by the hypoconid whose buccal base is somewhat widened apically. Entoconid low and elongated and joined to the distal hypoconid cristid. Weak basal cingulum. NM-Pv 11711, left m1 ( Text-fig. 6d View Text-fig 1–3 View Text-fig View Text-fig View Text-fig ) more slender but with similar morphology to that described above. NMPv 11712, left m1–m2, the m1 is somewhat smaller than the other carnassial teeth; the associated m2 is similar to the morphotype of NM-Pv 11713 described below.
NM-Pv 11747, left m1–m3 ( Text-fig. 6e View Text-fig 1–3 View Text-fig View Text-fig View Text-fig ); slightly smaller size compared to the rest of the specimens attributed to this species, m1 very robust, with a short paraconid, and a mesial cristid distally inclined. High and very robust protoconid. Metaconid still strong. Short talonid, almost
k1 k2 k3
completely occupied by the hypoconid, entoconid low and elongated and joined to the distal hypoconid cristid. The buccal basal wall is broken. The m2 is relatively large with respect to m1, trigonid with high protoconid somewhat larger than the metaconid, small mesiocentral paraconid. Talonid large with strong hypoconid attached to a peripherical entoconid. The m3 is well developed with a dominant protoconid, and vestigial metaconid and hypoconid. The talonid is robust.
NM-Pv 11713, left m2 ( Text-fig. 6f View Text-fig 1–3 View Text-fig View Text-fig View Text-fig ). It has a relatively high trigonid, dominated by the protoconid, although the metaconid is also quite well developed. The protoconid base is buccally widened. Very small paraconid, poorly differentiated from the anterior cristids of the protoconid and metaconid, which are joined together, closing mesially the trigonid valley. Talonid wide, with a high buccal hypoconid, clearly separated from the protoconid. The crestiform entoconid, low and peripheral, is joined distally to the distal hypoconid cristid, thus strongly delimiting the talonid valley, which is flat and very wide. Moderate basal cingulum, only strong in the mesiobuccal area. Another two m2, NM-Pv 11714 ( Text-fig. 6g View Text-fig 1–3 View Text-fig View Text-fig View Text-fig ) and NM-Pv 11712, display a similar morphology. NM-Pv 11718, right m2 ( Text-fig. 6h View Text-fig 1–4 View Text-fig View Text-fig View Text-fig View Text-fig ), NM-Pv 11716, left m2 ( Text-fig. 6i View Text-fig ) and Pv 11717, left m2 ( Text-fig. 6j View Text-fig 1–2 View Text-fig View Text-fig ) present a morphological pattern somewhat different from that of the teeth described above, in particular as a result of the narrow form of the talonid, which seems relatively more reduced.
NM-Pv 11696, is an association of a right m2 and m3 ( Text-fig. 6k View Text-fig 1–3 View Text-fig View Text-fig View Text-fig ). The m2 has a slightly better-developed paraconid than in the previously described specimens. The mesial wear facet with the m1 talonid is very clear. The m3 is oval in shape with a strong mesiobuccal protoconid, from which a mesial cristid extends to the lingual position, marking a small cuspid (paraconid). The talonid is poorly differentiated from the trigonid, and the hypoconid is very low and extends into a peripheral cristid that completely surrounds the talonid.
D i s c u s s i o n. Megamphicyon carnutense was defined by Antunes and Ginsburg (1977) as a new subspecies Amphicyon giganteus carnutense , in the same paper where they defined a new species Amphicyon olisiponensis, from the locality of Quinta do Narigao, Lisbon Basin, Portugal. According to these authors, Amphicyon olisiponensis appears to show affinities with both the mandible classified by Kuss (1965) as Pseudocyon sansaniensis aff. sansaniensis from Chilleurs, France, and the primitive forms of Amphicyon giganteus . Antunes and Ginsburg (1977) highlighted the difficulties involved in distinguishing between the Chilleurs form and Amphicyon olisiponensis. However, the small morphological differences between these two forms, together with the more modern age of the Lisbon site, lead them to maintain a specific distinction between the two. Therefore, according to these authors the Chilleurs form should be classified as Amphicyon giganteus , but differences in size lead them to propose a new subspecies A. giganteus carnutense .
Ginsburg (1989) added a third subspecies, Amphicyon giganteus laugnacensis, to distinguish the maxilla from Laugnac, determined by de Bonis (1973) as Amphicyon cf. astrei KUSS, 1962 from the other subspecies. He concluded that the Amphicyon giganteus species would comprise three successive stratigraphic subspecies; Amphicyon giganteus laugnacensis: MN 2; Amphicyon giganteus carnutense: MN 3; Amphicyon giganteus giganteus: MN 4a (Artenay), MN 4b (Baigneaux-en-Beauce), MN 5 (Pontlevoy, Falun d’Anjou). However, Ginsburg et al. (2000) subsequently reconsidered the taxonomic attribution of the Chilleurs mandible (type of Amphicyon giganteus carnutense ), considering the determination by Kuss to be correct (1965). Hence, the validity of Amphicyon giganteus carnutense was discarded. Therefore, the materials from Les Beilleaux attributed by Ginsburg (1989) to this taxon remained unnamed, and he proposed the new species Amphicyon (Megamphicyon) lathanicus for this fossil considering Megamphicyon KUSS, 1965 as a valid subgenus.
Nonetheless, we recognised many problems in the use of the denomination of Pseudocyon sansaniensis for the mandible from Chilleurs ( Kuss 1965: fig. 77). The Chilleurs species possesses a more robust m1 than that of the mandible type of P. sansaniensis ( Ginsburg 1961) but above all, it presents a significantly larger m2 relative to m1, in contrast with the small size of the m2 of the Sansan species. Thus, the decision of Antunes and Ginsburg (1977) remains valid; that the Chilleurs form and Amphicyon olisiponensis are very similar. Likewise, Ginsburg (1989) considered that A. olisiponensis could belong to the same group as Amphicyon giganteus . We think that the least confusing taxonomic hypothesis is that of Antunes and Ginsburg (1977), considering that the large Chilleurs form determined by Kuss (1965) as Pseudocyon sansaniensis should be called Megamphicyon carnutense , whilst Amphicyon (Megamphicyon) lathanicus should currently be considered as a synonym for the previous species, an opinion already expressed by Jiangzuo et al. (2019).
The dentition from Tuchořice is similar to that of Megamphicyon carnutense , but some teeth manifest morphological variations that may suggest the presence of a second species; as is the case of three m2 (specimens NMPv 11718, NM-Pv 11716 and NM-Pv 11717), which possess a narrow talonid, and are slightly smaller than the specimens with a subquadrate talonid. However, similar differences are found in the m2 of the Falun d’Anjou described as Amphicyon lathanicus by Ginsburg (2000b), in which morphotypes with a wide and subquadrate talonid (m2, FS 6953) coexist with others possessing a narrower talonid (FS 6965). We therefore regard all described specimens as belonging to a single species, Megamphicyon carnutense .




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Megamphicyon carnutense ( ANTUNES et GINSBURG, 1977)
| Morales, Jorge, Fejfar, Oldřich, Heizmann, Elmar, Wagner, Jan, Valenciano, Alberto & Abella, Juan 2021 |
Amphicyon (Megamphicyon) lathanicus
| Morales & Fejfar & Heizmann & Wagner & Valenciano & Abella 2021 |
Amphicyon giganteus carnutense
| Antunes et Ginsburg 1977 |