
Chromatophotina Rivera
|
publication ID |
https://doi.org/ 10.5281/zenodo.294122 |
|
DOI |
https://doi.org/10.5281/zenodo.6206549 |
|
persistent identifier |
https://treatment.plazi.org/id/038287CE-485B-FFA0-FF41-FDE4FED2FDB6 |
|
treatment provided by |
Plazi |
|
scientific name |
Chromatophotina Rivera |
| status |
|
Chromatophotina Rivera , new genus
Type species: Chromatophotina awajun , n. sp., by present designation.
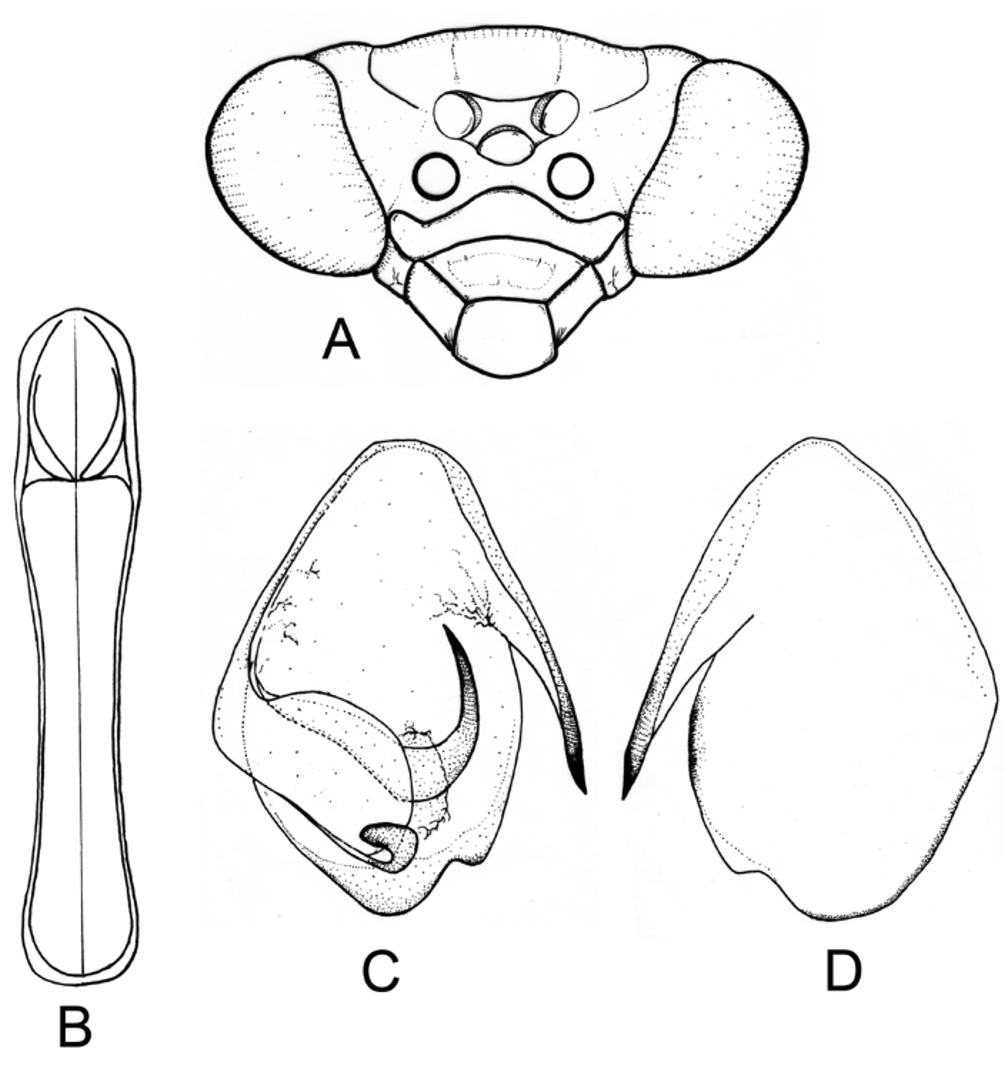
Description. Head pentagonal in shape; juxtaocular tubercles and vertex almost at the same level, forming a continuous edge, making the vertex slightly convex; frontal shield transversal, width approximately 4.6–6 times greater than length. Pronotum slender, supracoxal dilation weakly developed; lateral edges of the prozone almost parallel. Front femora with 3 discoidal and 5-6 external spines. Male: meso- and metathoracic wings well developed, hyaline and colorless, with iridescent reflections. Female: mesothoracic wing opaque; costal area covered with a white, dusty pigmentation, remaining of wing green; metathoracic wings squarish, anal area opaque, brightly colored from yellow to orange. Cerci 14-segmented. Male genitalia: ventral phallomere with an elongated and pointed lateral process; anterior process of the left dorsal phallomere curved and simple.
Etymology. The genus name is a combination of Chromato (from the greek Chr ō matikos, meaning “coloured”) and Photina , the type genus of the subfamily. The name makes reference to the intensely colored metathoracic wings of the females, unique among the Photinainae . The gender is female.
Systematics. Chromatophotina shows a number of morphological characters that clearly distinguish it from other photinaine genera. The combination of 5-6ES/3DS in the fore femora distinguishes this new genus from related taxa with similar spination formula, such as Hicetia Saussure and Zehntner, 1894 (6ES/4DS) and Microphotina Beier, 1935 (5ES/4DS). The number of discoidal and external spines of the fore femora is normally used as a diagnostic character in mantid taxonomy at different hierarchical levels. However, the discoidal spine character seems to be more reliable at the genus level because it is well conserved, unlike the number of external spines, which can show variability. For instance, the two new species of Chromatophotina described here have three discoidal spines but five to six external spines. The photinaine genus Macromantis is another good example of this variation, as it has four discoidal spines and from four to eight external (but tipically five to six) spines ( Roy 2002). This variable condition in the number of external spines also has been documented in other non-photinaine genera (i.e. Acanthops Serville, 1831 , Metilia Stål, 1877 , Stenophylla Westwood, 1843 ). Thus, I regard the presence of three discoidal spines as diagnostic for Chromatophotina but it is by no means an autopomorphy as other unrelated Photinainae genera share this feature.
The presence of five to six external and three discoidal spines in the fore femora, and particularly the white coloration of the costal area of the mesothoracic wings and the strikingly colored metathoracic wings in the female, allow this genus to be separated from other Photinainae . Pigmentation involving the whole surface of the metathoracic wings is rare in Photinainae , and if present, is restricted to dull brown, as in females of Orthoderella , some species of Coptopteryx with brown color morphs (unpublished observation) and Paraphotina ( Cerdá 1992) . In other Photinainae , green pigmentation on the metathoracic wings may be restricted to a small area immediately surrounding the cross-veins on the anal field, leaving most of the wing surface hyaline and colorless. The bright and stunning pigmentation of the metathoracic wings of Chromatophotina makes this new taxon unique among the Photinainae .
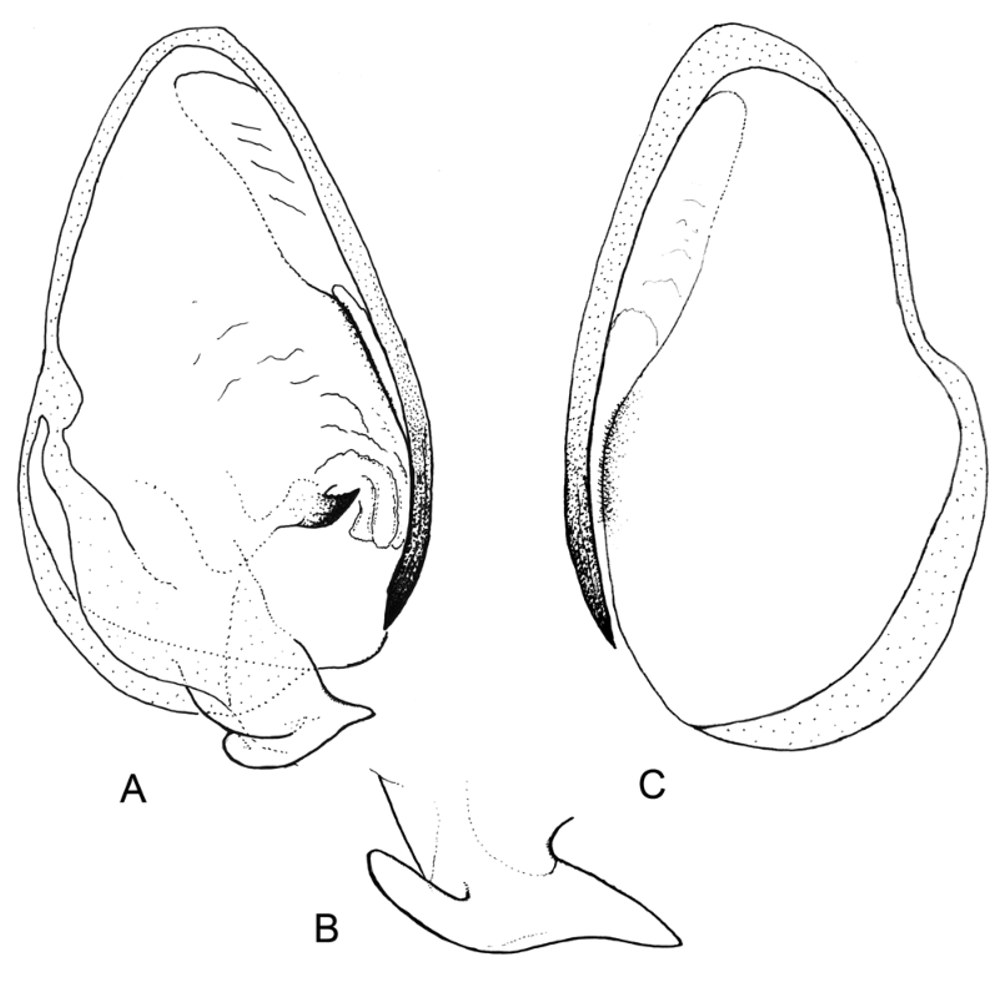
The examination of the male genitalia allowed assignment of Chromatophotina to the Photinaini (as currently conceived) and near to Microphotina . Both genera share the elongated and pointed lateral process of the ventral phallomere and a well developed anterior process of the left dorsal phallomere, which in Chromatophotina is simple ( Fig. 2 View FIGURE 2 C,D) and in Microphotina forked and hammerhead-like ( Fig. 5 View FIGURE 5 A, B; Cerdá 1997, Fig. 94). On the other hand, the genus Hicetia , which is usually cited in the literature as related to Microphotina ( Ehrmann, 2002) , seems not to be closely related to this latter genus or Chromatophotina . The male genitalia of Hicetia goeldiana Saussure and Zehntner, 1894 was studied and illustrated by Heitzmann (1965) (under its synonym Metriomantis gracilicollis Beier, 1935 ). The morphology of the male genitalia in Hicetia shows more similarities to that of other Photinaini genera such as Photina Burmeister, 1838 and Photinella Giglio-Tos, 1915 , where they all share a characteristic short, voluminous and recurved lateral process on the ventral phallomere (as opposed to slender, sharp and elongated as in Microphotina and Chromatophotina ).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.