
Coelodonta antiquitatis (Blumenbach, 1799)
|
publication ID |
https://doi.org/10.5252/g2015n2a5 |
|
publication LSID |
urn:lsid:zoobank.org:pub:0117CBA4-4CE0-4431-B5F6-721F998C72C7 |
|
persistent identifier |
https://treatment.plazi.org/id/038087D7-FFFB-FF80-FC97-FA9DFDF530E8 |
|
treatment provided by |
Felipe |
|
scientific name |
Coelodonta antiquitatis (Blumenbach, 1799) |
| status |
|
Coelodonta antiquitatis (Blumenbach, 1799)
MATERIEL EXAMINED. — NISP=225; MNI=39.
1842 sample: 1 right M2; 3 tooth fragments; 1 right humerus; 1 left humerus; 1 left ulna; 1 right innominate; 1 innominate; 1 right tibia; 1 right talus.
1989-1992 sample: 2 petrosal bones; 4 left D1; 4 right D2; 4 left D2; 12 right D3; 8 left D3; 5 right D4; 7 left D4; 1 upper deciduous tooth fragment; 1 right P2; 3 left P2; 2 right P3; 1 left P3; 2 right P4; 1 left P4; 1 right M1; 4 left M1; 1 right M2; 4 left M2; 3 right M3; 2 left M3; 2 upper molar fragments; 2 right d1; 1 left d1; 4 left d2; 3 right d3; 9 left d3; 6 right d4; 1 left d4; 1 d4; 2 lower deciduous tooth fragments; 1 left p2; 1 p2-3; 2 right p3; 1 right p3-4; 3 left p3-4; 4 lower premolar fragments; 4 right m1; 1 left m1; 1 left m2-3; 1 right m3; 2 left m3; 2 lower molar fragments; 55 tooth fragments; 1 left scapula; 2 right humeri; 2 left humeri; 2 radii; 1 left metacarpal IV; 2 left innominates; 1 right femur; 1 femur; 4 right tibiae; 4 left tibiae; 6 tibiae; 1 right talus; 1 first phalanx; 3 long bone fragments; 1 flat bone fragment (scapula or innominate); 1 metapodial.
DESCRIPTION
More than 90 isolated teeth (mainly deciduous teeth), and several postcranial elements have been attributed to the woolly rhinoceros C. antiquitatis (Blumenbach, 1799) ( Fig.8 View FIG ). The morphology of this Eurasian Pleistocene species is well-known (for review, see Guérin 1980). The Fouvent sample presents all the morphological features of this species (large size, high crowned teeth, quadrangular M3s with distinct ectoloph and metaloph).
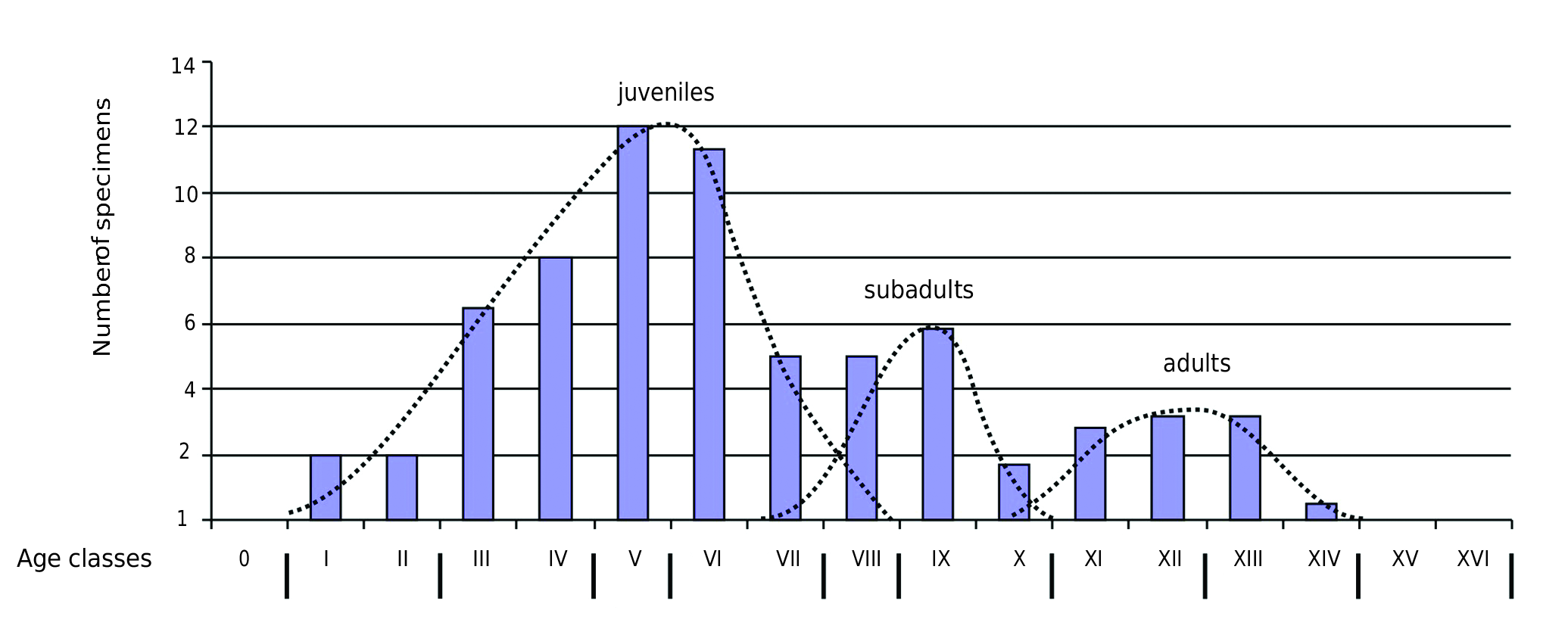
We have used the tooth wear stage-based protocol developed by one of us (POA) for reconstructing mortality curves in both extant and extinct rhinoceroses, based on isolated teeth ( Bacon et al. 2008). The age classes as characterized on the white rhinoceros by Hillman-Smith et al. (1986) are used in this study because of the phylogenetic, chronological and ecological close relationships between this extant species and the woolly species ( Antoine 2002). We have restricted this analysis to upper teeth, more abundant in Fouvent ( 69 specimens), as they display a much more homogenous eruption and wear pattern than the lower teeth, both in the white rhino and in the woolly rhino. The mortality curve obtained for C. antiquitatis is trimodal, with a majority of individuals ranging from classes I to IX (1.5 months up to 9 years) ( Fig. 10 View FIG ); the first mode is comprised between 1.5 and 4 years (classes V-VI), which coincides with a period encompassing weaning and abandonment of juvenile individuals by the mother ( Groves 1972). The second mode includes specimens the individual age of which ranges from 4 and 9 years, e.g., more or less the subadult-adult transition (classes VIII-IX; Hillman-Smith et al. 1986). The third and last mode, with less specimens, includes adult individuals (14-28 years; classes XII-XIII). Based on upper teeth, juveniles (61%) are far overrepresented with respect to subadults (23%) and adults (16%). Such a structure is significantly distinct from that of natural populations of recent Indian rhinos ( Rhinoceros unicornis Linnaeus, 1758), consisting of 27% juveniles, of 21% subadults, and of 52% adults ( Laurie 1982; Laurie et al. 1983). On the other hand, the Fouvent mortality profile is much similar to what is observed for the late Pleistocene rhino tooth sample of Duoi U’Oi, in Vietnam and other Southeast Asian cave localities, for which the accumulating factor is also of biological origin (porcupines; Bacon et al. 2008; Antoine 2012). Nevertheless, 14 woolly rhino teeth from the 1989-1992 excavation sample in Fouvent are eroded, which attests to a by-pass and a hydrodynamical erosion, both post-mortem and pre-accumulation (n°32, 42, 78, 87, 107, 124, 158, 192, 225, 269, 293, 330, 337, and 480). A lower tooth was split into two pieces then eroded, before both halves were recovered from two distinct excavation units (FVT 92 G9B-525 and FVT 92 F9B-165).
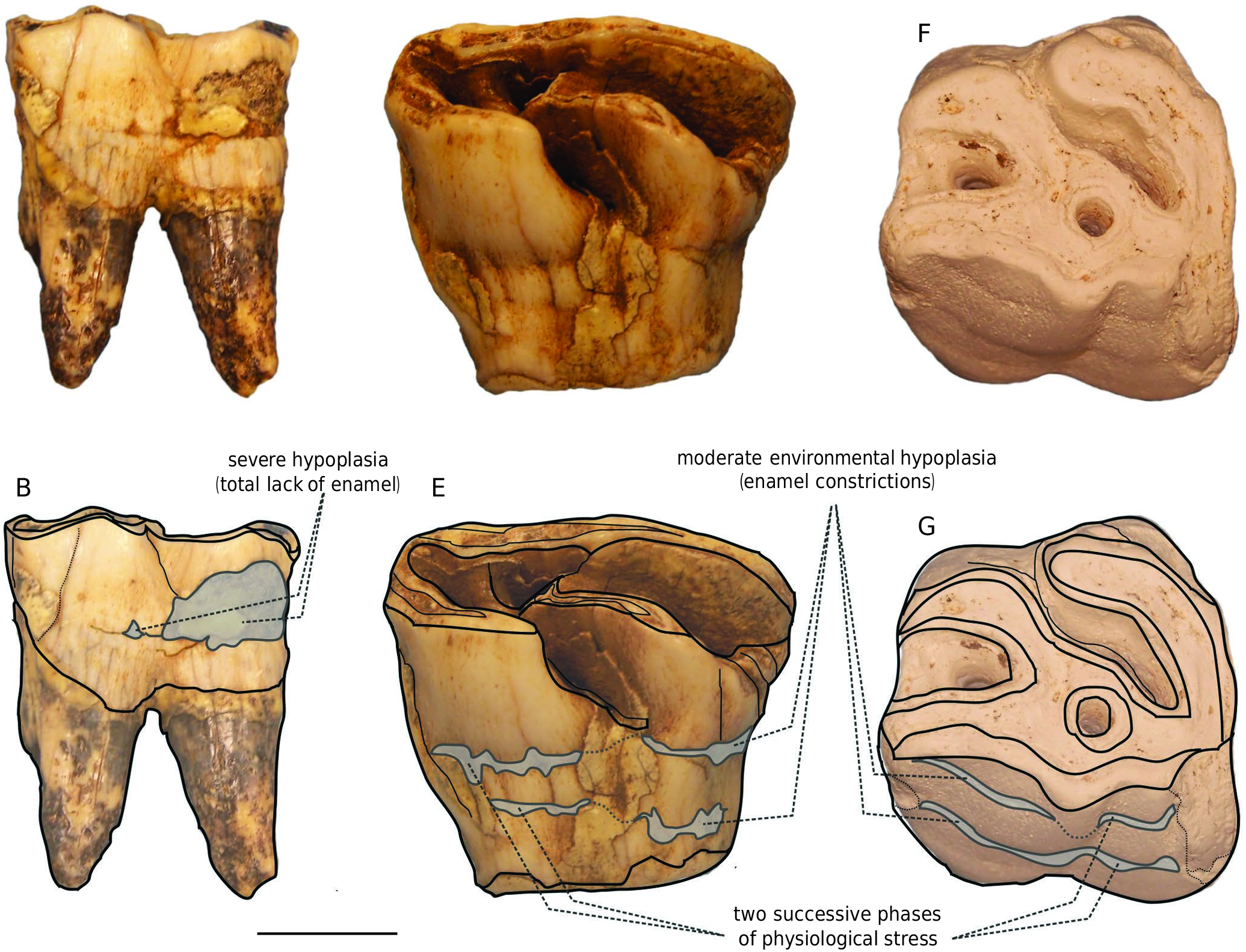
Several teeth from Fouvent show environmental enamel hypoplasia, which expresses a severe physiological stress during odontogenesis (but does not imply any genetic origin). On decidual teeth, this pathology is due to a starving episode for the mother during pregnancy or nursing, whereas enamel hypoplasia on permanent teeth expresses denutrition coinciding with either weaning or abandonment of the calf by a newly pregnant mother ( Mead 1999). In Fouvent, the specimens showing the most spectacular pathologies are the M1 G9D 607, the M2 E11B 218, and the d4 F11 C243 ( Fig. 11 View FIG ).
Individual ages 1.5 month 4 months 1.5 year 3 years 4 years 7 years 10-11 years 20 years 30-32 years 40 years
Order ARTIODACTYLA Owen, 1848 Suborder RUMINANTIA Scopoli, 1777 Family BOVIDAE Gray, 1821 Subfamily BOVINAE Gray, 1821 Genus Bison Smith, 1827

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.