
Apocellus andinus, Chani-Posse, Mariana & Scheibler, Erica E., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3721.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:9FB5C00A-01B4-4CFD-BDD6-6556E9641A01 |
|
DOI |
https://doi.org/10.5281/zenodo.6158004 |
|
persistent identifier |
https://treatment.plazi.org/id/F6283D31-8F7A-B72F-E1D5-7270C0E6FDCF |
|
treatment provided by |
Plazi |
|
scientific name |
Apocellus andinus |
| status |
sp. nov. |
Apocellus andinus View in CoL sp. nov.
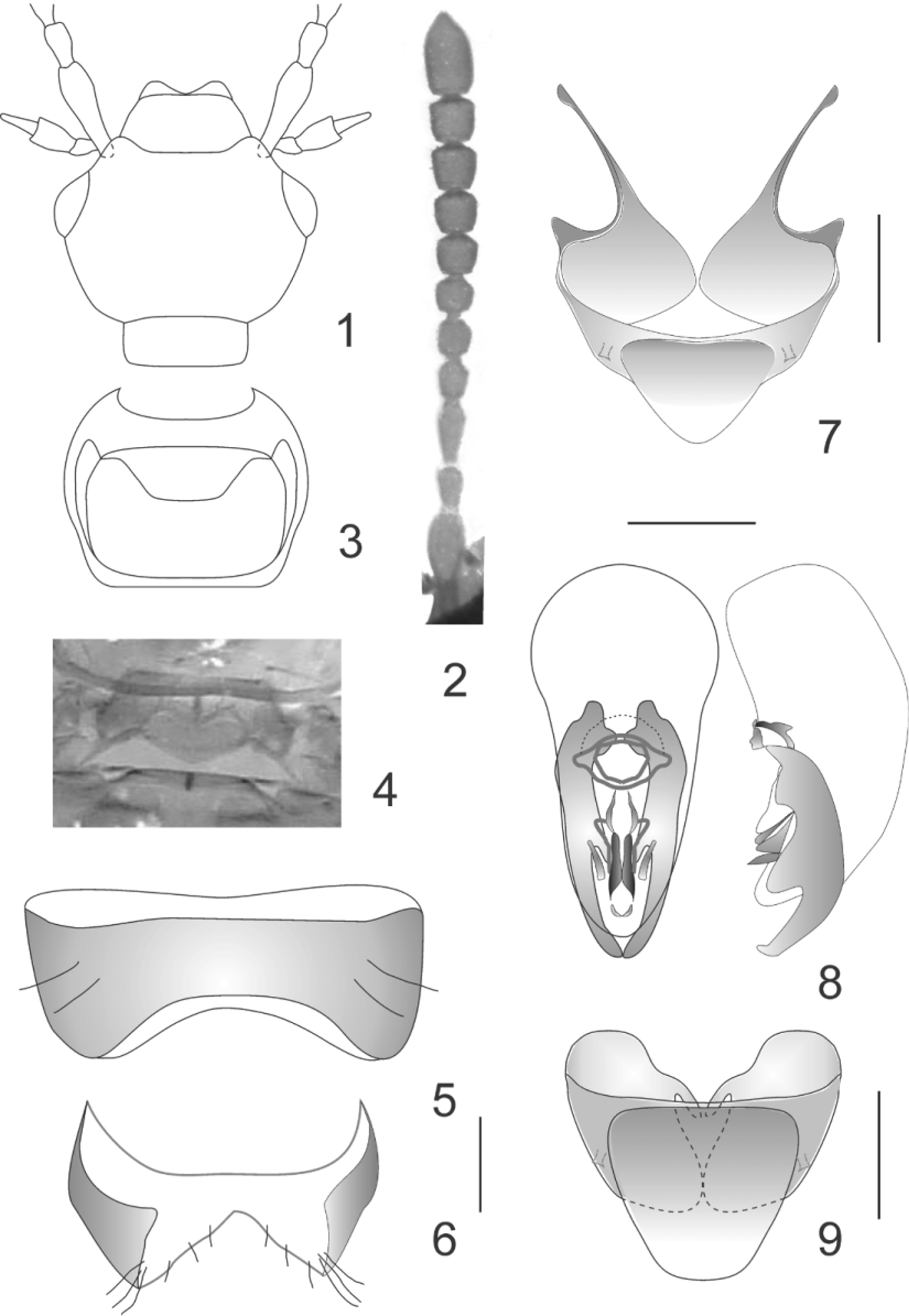
( Figs 1–9 View FIGURES 1 – 9 )
Diagnosis. Apocellus andinus is very similar in general appearance to the southern South American species A. mendozanus Steinheil. Both species have eyes as long as the temples seen from above and the neck is about one half as wide as the head behind the eyes. The new species may be identified, and distinguished from A. mendozanus , by having the head slightly narrowed behind the eyes and the antennal segments 7 to 10 quadrate.
Description. Body length 3.5–4.0 mm, body shape somewhat myrmecoid, head and thorax distinctly narrower than elytra and abdomen; pronotum subrectangular and globose; abdomen tapering toward fifth and sixth visible abdominal terga, with distinctly scarce setation. Coloration. Head, thorax, elytra and abdomen testaceousbrunneous, apical (sixth to eight visible) abdominal segments darker; antennae, palpi and legs testaceous.
Head subrectangular, slightly narrowed behind eyes, with broadly rounded hind angles ( Fig. 1 View FIGURES 1 – 9 ); moderately wider than long (HW/HL= 1.30–1.35), about as wide as to slightly wider than pronotum at widest point (HW/PW= 1.05–1.07); dorsal and ventral surface of head glabrous with fine and sparse punctuation; labrum, clypeus and frons each with one pair of short and strong setae at the anterior margin. Eyes distinctly convex and coarsely facetted, as long as temples (EL/TL= 1.0) seen from above ( Fig. 1 View FIGURES 1 – 9 ). Antennae inserted on ridges at epistomal suture ( Fig. 1 View FIGURES 1 – 9 ), moderately long, filiform; first three segments bearing only sparse setae, segments 4 to 11 densely punctuate, segment 3 longer than segment 2, segments 2 + 3 about as long as to slightly longer than segment 1, segment 4 elongate, segments 5 to 10 quadrate, last segment about twice as long as penultimate segment and narrowed into a subacute apex ( Fig. 2 View FIGURES 1 – 9 ). Labrum transverse and broadly arcuate ( Fig. 1 View FIGURES 1 – 9 ). Clypeus well developed, subrectangular. Epistomal suture distinct ( Fig. 1 View FIGURES 1 – 9 ). Mandibles stout and symmetric, each with four teeth. Maxilla with galea elongate and densely setose at apex, galea much shorter than lacinia and densely setose medially. Maxillary palpus with last segment acicular and about one half as long as segment 3 ( Fig. 1 View FIGURES 1 – 9 ), segment 3 robust and broadened apically, segment 2 narrower than maximum width of segment 3 and no more than 0.7 times as long, segment 1 minute. Gular sutures confluent on anterior half and strongly divergent near base of head. Mentum and submentum subequal in length. Labial palpus with first two segments subequal in length, segment 3 slightly shorter and narrower than segment 2. Neck one half as wide as head at widest point (NW/HW= 0.5).
Pronotum as wide as to slightly wider than long (PW/PL= 1.05–1.12), narrowed posteriad; front margin broadly arcuate, hind margin truncate, anterior angles broadly rounded, posterior angles obtusely rounded; surface with fine and sparse punctuation; hypomeron broad and slightly inflexed (visible in lateral view of prothorax). Prosternum subtrapezoidal ( Fig. 3 View FIGURES 1 – 9 ). Mesosternum with mesosternal process short and truncate apically. Scutellum small, triangular, with a heart-shaped impression ( Fig. 4 View FIGURES 1 – 9 ). Legs moderately long with three segmented tarsi; surface of all segments glabrous except for scattered, short marginal setae; front femora as long as front tibiae; all tarsi with first and second segments reduced and subequal in length, third tarsal segment one third as long as tibia, tarsal claws distinctly developed, about one half as long as third tarsal segment.
Elytral epipleural ridge present. Elytra both at suture (Etl/PL=1.05–1.12) and at sides slightly longer than pronotum at midline (EtL/PL=1.15–1.20), at base wider than pronotum; punctuation fine and scattered.
Abdomen. Terga 2 to 7 (first to sixth visible) with basolateral ridge; tergum 7 (sixth visible) with whitish apical seam of microtrichae; surface with scattered minute punctuation. Tergum 8 (seventh visible) with posterior margin subtruncate.
Male. Sternum 7 with posterior margin broadly and deeply arcuate and membranous ( Fig. 5 View FIGURES 1 – 9 ). Sternum 8 with posterior margin broadly and deeply emarginate, strongly reduced at middle and mostly membranous, excepting lateral margins ( Fig. 6 View FIGURES 1 – 9 ). Tergum 9 with two pairs of ventral struts, the anteromedial pair distinctly longer than the anterolateral pair ( Fig. 7 View FIGURES 1 – 9 ). Tergum 10 with posterior margin subangulate ( Fig. 7 View FIGURES 1 – 9 ); slightly sclerotized apically, with two long subapical macrosetae and two short and fine apical setae. Aedeagus with parameres moderately long and broad, branched at middle, converging apically beyond apex of median lobe. Median lobe with base bulbous and apical portion truncate. Internal armature of median lobe well sclerotized, with two pairs of short longitudinal processes ( Fig. 8 View FIGURES 1 – 9 ).
Female. Tergum 9 with one pair of medioventral struts ( Fig. 9 View FIGURES 1 – 9 ). Tergum 10 with posterior margin subtruncate ( Fig. 9 View FIGURES 1 – 9 ). Spermatheca with receptacle sclerotized.
Etymology. The specific name andinus refers to the Andes where the species was discovered.
Type material. Holotype, ♂, with labels: “ Argentina. Mendoza: Dto. Luján de Cuyo, Potrerillos, S 32º 57.335' W 69º 10.840', 15-II-2008, wetland, 1350 m, E. Scheibler”, “ Holotype Apocellus andinus n. sp. des. Chani-Posse & Scheibler, 2013” (IADIZA). Allotype, ♀, with the same data as holotype (IADIZA). Four paratypes with the same data as holotype (2 IADIZA, 2 FMNH).
Distribution. Known only from the type locality (Andean wetland at 1350 m elevation) in northwestern Mendoza Province, Argentina.
Apocellus andinus n. sp. was found in a riparian zone from the Potrerillos dam vulnerable to periods of drought depending on the dam water fluctuations. During the wet phases, the system exhibited highest values of conductivity (Mean= 895.17 µS cm -1; SD = 300.45), neutral pH values (Mean= 7.29; SD= 0.29) and temperate waters (Mean= 16.85 ºC, SD= 4.14). In the sampling period, water temperature varied between maximal values corresponding to the month of February (23.3 ºC) and minimal values on July (11.2 ºC). The percentage of humidity changed between 28% and 74% during the sampling period. Wetland depth reached minimal values of 0.10 m and maximal values of 0.61 m and the percentage of oxygen saturation had mean values of 78.37%.
Specimens of Apocellus andinus were found during February where the environmental conditions showed the highest values of air and water temperature (28.7ºC and 23.3ºC respectively), great depth (0.50 m) as well as high values of water mineralization (696 µS cm -1) and oxygenation (77% of oxygen saturation). The substrate was composed mainly by sand and silt. The vegetation was represented by Poplar trees and algae ( Cladophora sp.) to a lesser extent in the sampling site ( Figs. 13, 14 View FIGURES 13 – 14 ).
Bionomics. Apocellus andinus n. sp. was collected in association with species of Sigara (Tropocorixa) jensenhaarupi Jaczewski ( Heteroptera , Corixidae ) and Berosus sp. ( Coleoptera , Hydrophilidae ). Sigara (T.) jensenhaarupi is endemic to Argentina and it is widely distributed in both Andean and extra-Andean environments from Mendoza province (Melo and Scheibler, 2011). This species can tolerate a wide range of salinity in permanent and temporary waters with either scarce or abundant riparian vegetation in extremely arid regions (Bachmann, 1981; Muzón et al., 2005; Melo and Scheibler, 2011; Scheibler and Ciocco, 2011) which are highly impacted by human activities (Muzón et al., 2005). On the other hand, Berosus species are well known by their tolerance to diverse pollution sources and they have shown to be largely indifferent to organic pollution (Hellawell, 1986). Species belonging to this genus were found in pools highly exposed to human pollution (von Ellenrieder and Fernández, 2000) as well as in hard and saline waters of the Llancanelo lake (Scheibler and Ciocco, 2011), a Ramsar wetland located in the south of Mendoza province.
Among others invertebrates associated with Apocellus andinus , we can mention the following taxa: Hyalella sp. (Anfipoda, Crustacea), Lumbriculidae sp. (Oligochaeta), Chironomidae (Diptera) , Rhionaeschna absoluta Calvert ( Odonata : Aeshnidae ). Cyanallagma interruptum Selys ( Odonata : Coenagrionidae ), Tropisternus lateralis (Say) ( Coleoptera : Hydrophilidae ) and Liodessus flavofasciatus (Steinheil) ( Coleoptera : Dytiscidae ). Cyanallagma interruptum and Rhionaeschna absoluta had been collected in temporary ponds of areas impacted by cattle and modified by human action. (Muzón et al., 2005). Tropisternus lateralis , on the other hand, does not show drought resistance such as other invertebrates do (e. g. Chironomidae larvae, some Crustacea) and it tends to quickly colonize non-permanent or temporary habitats (e.g. cattle drinking troughs) (Cook and Kennedy, 2000).
Apocellus andinus n. sp. was found once in the unique non-permanent wetland of five lentic sampling sites chosen for a spatial and temporal study of the aquatic entomofauna in the northwest of Mendoza province. Due to the unstable conditions of the area where A. andinus was found and the type of association that it had with other macroinvertebrates, we infer that this species belongs to a transient fauna (Scheibler and Ciocco, 2011) adapted to dry periods. From its association with Berosus sp., Sigara (T.) jensenhaarupi , Cyanallagma interruptum and Rhionaeschna absoluta , and given the high conductivity values (800 µS cm -1 aprox.) of its system, A. andinus could be tolerant to saline waters and environmental impacts caused by human activities. However, further collecting effort must be done to increase the knowledge of the biology and distributional range of this new species.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |